An official website of the United States government
The .gov means it’s official.
Federal government websites often end in .gov or .mil. Before
sharing sensitive information, make sure you’re on a federal
government site.
The site is secure.
The https:// ensures that you are connecting to the
official website and that any information you provide is encrypted
and transmitted securely.
1 Translational Tissue Engineering Center, Wilmer Eye Institute and Department of Biomedical Engineering, Johns Hopkins University School of Medicine, Baltimore, Maryland, USA.
2 Department of Biomedical Sciences, Korea University College of Medicine, Seoul, South Korea.
3 Bloomberg-Kimmel Institute for Cancer Immunotherapy and.
4 Sidney Kimmel Comprehensive Cancer Center, Johns Hopkins University School of Medicine, Baltimore, Maryland, USA.
5 Buck Institute for Research on Aging, Novato, California, USA.
6 Department of Pharmacodynamics, College of Pharmacy, University of Florida, Gainesville, Florida, USA.
7 Division of Rheumatology, Johns Hopkins University School of Medicine, Baltimore, Maryland, USA.
1 Translational Tissue Engineering Center, Wilmer Eye Institute and Department of Biomedical Engineering, Johns Hopkins University School of Medicine, Baltimore, Maryland, USA.
2 Department of Biomedical Sciences, Korea University College of Medicine, Seoul, South Korea.
3 Bloomberg-Kimmel Institute for Cancer Immunotherapy and.
4 Sidney Kimmel Comprehensive Cancer Center, Johns Hopkins University School of Medicine, Baltimore, Maryland, USA.
5 Buck Institute for Research on Aging, Novato, California, USA.
6 Department of Pharmacodynamics, College of Pharmacy, University of Florida, Gainesville, Florida, USA.
7 Division of Rheumatology, Johns Hopkins University School of Medicine, Baltimore, Maryland, USA.
Senescent cells (SnCs) are implicated in the pathogenesis of age-related diseases including osteoarthritis (OA), in part via expression of a senescence-associated secretory phenotype (SASP) that includes immunologically relevant factors and cytokines. In a model of posttraumatic OA (PTOA), anterior cruciate ligament transection (ACLT) induced a type 17 immune response in the articular compartment and draining inguinal lymph nodes (LNs) that paralleled expression of the senescence marker p16INK4a (Cdkn2a) and p21 (Cdkn1a). Innate lymphoid cells, γδ+ T cells, and CD4+ T cells contributed to IL-17 expression. Intra-articular injection of IL-17-neutralizing antibody reduced joint degeneration and decreased expression of the senescence marker Cdkn1a. Local and systemic senolysis was required to attenuate tissue damage in aged animals and was associated with decreased IL-17 and increased IL-4 expression in the articular joint and draining LNs. In vitro, we found that Th17 cells induced senescence in fibroblasts and that SnCs skewed naive T cells toward Th17 or Th1, depending on the presence of TGF-β. The SASP profile of the inflammation-induced SnCs included altered Wnt signaling, tissue remodeling, and cell-cycle pathways not previously implicated in senescence. These findings provide molecular targets and mechanisms for senescence induction and therapeutic strategies to support tissue healing in an aged environment.
Conflict of interest: DZ is an inventor of a pending patent application for BCL-xL inhibitors and targeted senolytic agents for age-related diseases (Compounds that induce degradation of anti-apoptotic BCL-2 family proteins and the uses thereof, no. US20190135801). DZ and JC are cofounders, advisors for, and shareholders of Unity Biotechnology, which develops senolytic agents. JHE is an equity holder of Unity Biotechnology, receives patent royalties through Johns Hopkins University School of Medicine (Unit dose of a cis-imidazoline for treating an osteoarthritic joint by removing senescent cells, no. US9849128B2), and is a founder and equity holder of Aegeria Soft Tissue (AST). DMP is a consultant for Aduro Biotech, Amgen, Astra Zeneca, Bayer, DNAtrix, Dynavax Technologies, Ervaxx, FLX Bio, Janssen, Merck, and Rock Springs Capital. DMP also provides research support and consulting to Compugen and Bristol Myers Squibb and receives patent royalties through Johns Hopkins University School of Medicine. DMP is on the scientific advisory board of Camden Nexus II; is on the board of directors of and owns stock in Dracen Pharmaceuticals, Five Prime Therapeutics, and WindMIL Therapeutics; is a consultant for Immunomic Therapeutics and Tizona Therapeutics; has equity and patent royalties through Johns Hopkins University School of Medicine (Use of multivalent chimeric peptide-loaded, MHC/lg molecules to detect, activate or suppress antigen-specific, T cell-dependent immune responses; no. PCT/US98/18909; Manipulation of regulatory T cells using LAG-3 antibodies, genes, agonists and antagonists, no. PCT/US04/006133; Microsatellite instability as a pharmacogenomic marker of therapeutic response to immune checkpoint inhibition, no. PCT/US2015/060331; Immune checkpoint chimeric antigen receptors (CAR) therapy, no. PCT/US2016/040010; Marrow infiltrating lymphocytes (MILs) as a source of T-cells for chimeric antigen receptor (CAR) therapy, no. PCT/US2016/041521; Combination of immunotherapy with local chemotherapy for the treatment of malignancies, no. PCT/US2016/028861; Use of SON-211 and Ontak as a regulatory T cell targeted cancer immunotherapy in combination with anti-PD-1 therapy for the treatment of cancer, no. PCT/US2019/020959); and has founder equity in Potenza Therapeutics and Trieza Therapeutics.
Figures
Figure 1. Adaptive immune cells respond to…
Figure 1. Adaptive immune cells respond to traumatic joint injury with a type 17 immune…
Figure 1. Adaptive immune cells respond to traumatic joint injury with a type 17 immune response.
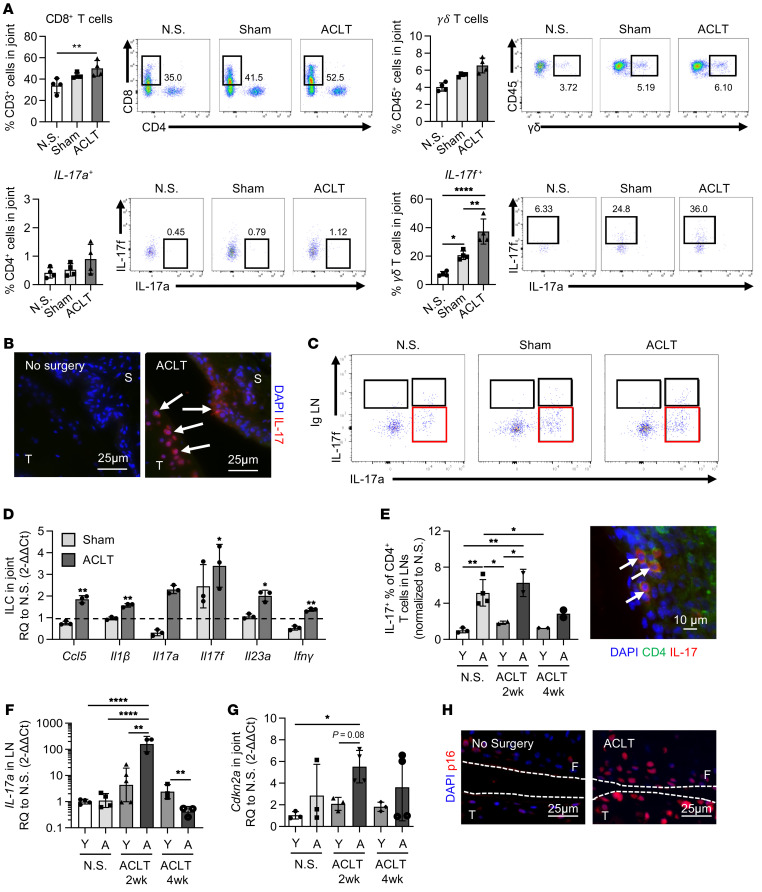
(A) Multiparametric flow cytometric analysis of CD8+, CD4+, and yδ+ T cells (CD45+CD3+) isolated from the joint compartment 1 week after sham surgery and ACLT compared with control mice with no surgery (N.S.) (n = 4). (B) Immunofluorescence of IL-17 in the synovium and cartilage 1 week after ACLT compared with no surgery in young mice. Scale bars: 25 μm. (C) Flow cytometric data showing IL-17a and IL-17f expression in ILCs from inguinal (Ig) LNs 4 weeks after ACLT (CD3–Thy1.2+NK1.1–). (D) Quantification of mRNA expression for inflammatory markers in ILCs (CD3–Thy1.2+) sorted from the joint compartment 2 weeks after ACLT (n = 2). (E) Percentage of Th17 cells in young (Y) and 18-month-old aged (A) animals 2 and 4 weeks after ACLT in the inguinal LNs, as determined by flow cytometry and immunofluorescence staining for CD4 and IL-17 in LNs from young mice 2 weeks after ACLT. Scale bar: 10 μm. (F) Quantification of Il17 mRNA expression in LN tissue (n = 3). (G) Quantification of Cdkn2a mRNA expression in young and aged animal joints with no surgery and in joints 2 and 4 weeks after ACLT (n = 3). (H) p16 staining of ACLT cartilage and no-surgery cartilage from young mice. Scale bars: 25 μm. Data indicate the mean ± SD. *P < 0.05, **P < 0.01, and ****P < 0.001, by 1-way ANOVA with Holm-Šidák multiple-comparisons test. All groups were compared with each other. (E–G) Separate 1-way ANOVAs were performed for each time point. F, femur; S, synovium; T, tibia; RQ, relative quantification.
Figure 2. Clearance of SnCs reduces the…
Figure 2. Clearance of SnCs reduces the IL-17 immune signature.
( A ) Quantification of …
Figure 2. Clearance of SnCs reduces the IL-17 immune signature.
(A) Quantification of Cdkn2a mRNA expression in articular joints of young and aged mice that had no surgery (N.S.), mice that underwent sham surgery, and mice that underwent ACLT and treatment with vehicle (Veh), i.a. UBX0101 (UBX), i.a. UBX plus i.p. Navi, or i.p. Navi (n = 3–6). (B) Quantification of Il17f mRNA expression in articular joints of young and aged mice as in A (n = 3–6). (C) Percentage of weight placed on the operated limb versus the contralateral control limb by in mice as in A (n = 3–6). (D) Quantification of CD4+IL-17a+ cells from inguinal LNs of mice and representative flow plots (n = 3 for each group). (E) Quantification of mRNA expression for Il17a in inguinal LN tissue (n = 3 for each group). (F) Representative safranin-O images of joints and OARSI scores for mice as in A (n = 3–6). Original magnification, ×20. (G) Representative safranin-O images of joints and OARSI scores after ACLT and treatment with isotype or IL-17a NAb ( n = 3). Original magnification, ×20. (H) Percentage of weight placed on the operated limb versus the contralateral control limb for mice that had no surgery or mice after ACLT that were treated with either the isotype control or a IL-17a NAb (n = 9). (I) Quantification of Cdkn1a mRNA expression in articular joints (n = 3). Data indicate the mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.005, and ****P < 0.001, by 1-way ANOVA with Holm-Šidák multiple-comparisons test. All groups were compared with each other. (A–F) Separate 1-way ANOVAs were performed for young and aged groups, and experimental groups were compared with the control group (vehicle-treated mice).
Figure 3. Inflammation-induced senescence in fibroblasts cultured…
Figure 3. Inflammation-induced senescence in fibroblasts cultured with Th17 cells and T cell skewing by…
Figure 3. Inflammation-induced senescence in fibroblasts cultured with Th17 cells and T cell skewing by senescent fibroblasts.
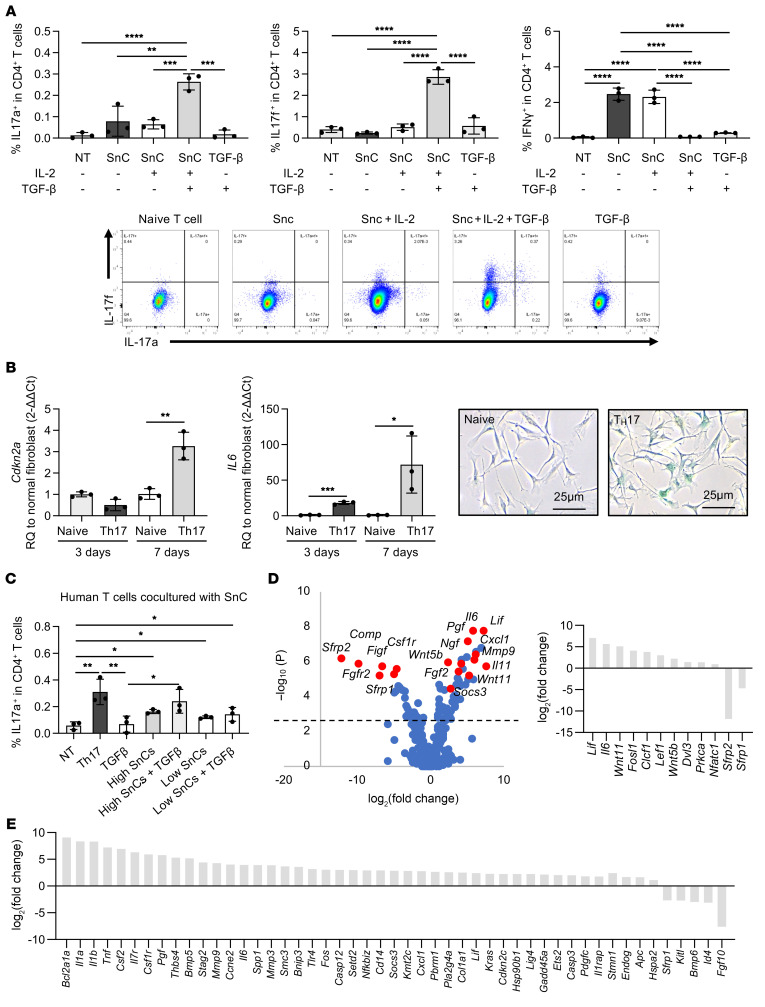
(A) Percentage of mouse CD4+IL-17a+, CD4+IL-17f+, and CD4+IFN-γ+ T cells in coculture conditions: naive T cells alone (NT), SnC coculture, SnCs plus IL-2 plus TGF-β coculture, media with TGF-β (n = 3 for each group). Representative flow plots are shown. (B) Quantification of Cdkn2a and Il6 mRNA expression in fibroblasts cocultured with naive CD4+ T cells and Th17 cells (n = 3 for each group). Representative images of SA–β-gal staining of fibroblasts cocultured with naive CD4+ T cells and Th17 cells are shown. Scale bars: 25 μm. (C) Percentage of human CD4+IL-17a+ T cells after 7 days in coculture. Conditions from left to right: no coculture (NT), Th17 positive control, high percentage of SnCs coculture, high percentage of SnCs plus IL-2 plus TGF-β coculture, low percentage of SnCs coculture, low percentage of SnCs plus IL-2 plus TGF-β coculture (n = 3 for each group). (D) Volcano plot of Th17 cocultured cells normalized to normal fibroblasts. The dotted line denotes P = 0.05. Graph on the right indicates differentially regulated genes related to STAT3 and Wnt signaling. (E) Differential expression of the top-50 genes in immune-induced senescence (Th17 coculture) compared with irradiation-induced senescence, 7 days after senescence induction (n = 3). Data indicate the mean ± SD. *P < 0.05 and **P < 0.01, by 1-way ANOVA with Holm-Šidák multiple-comparisons test. All groups were compared with each other. A 2-tailed Student’s t test was performed within each time point in B.
Figure 4. IL-4 is required for wound…
Figure 4. IL-4 is required for wound healing after SnC clearance and IL-17 reduction.
( …
Figure 4. IL-4 is required for wound healing after SnC clearance and IL-17 reduction.
(A) Quantification of Il4 mRNA expression in articular joints of young and aged mice (no surgery, sham surgery, after ACLT treated with vehicle, i.a. UBX, i.a. UBX plus i.p. Navi, and i.p. Navi). n = 3–5. (B) Quantification of Il4 mRNA expression in inguinal LNs (n = 3). (C) Percentage of CD4+IL-4+ T cells in inguinal LNs (n = 3). Representative flow plots of cells from LNs from young animals (no surgery, vehicle-treated, i.a. UBX–treated, and i.p. Navi–treated). (D) Percentage of weight placed on the operated limb versus the contralateral control limb (n = 4). (E) Gene expression of Il17a in inguinal LNs from WT versus Il4r–/– mice after treatment with senolytic drug (n = 3). (F) OARSI score of young WT and Il4r–/– joints treated with vehicle or senolytic drug. (G) Representative images of young WT or Il4r–/– joints treated with vehicle or senolytic drug. WT vehicle and i.a. UBX images are also shown in Figure 2F. Original magnification, ×20. Data indicate the mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.005, and ****P < 0.001, by 1-way ANOVA with Holm-Šidák multiple-comparisons test. Experimental groups were compared with the control group (vehicle).
Jeon OH, et al. Local clearance of senescent cells attenuates the development of post-traumatic osteoarthritis and creates a pro-regenerative environment. Nat Med. 2017;23(6):775–781. doi: 10.1038/nm.4324.
-
DOI
-
PMC
-
PubMed
Blau HM, Cosgrove BD, Ho AT. The central role of muscle stem cells in regenerative failure with aging. Nat Med. 2015;21(8):854–862. doi: 10.1038/nm.3918.
-
DOI
-
PMC
-
PubMed
Bussian TJ, Aziz A, Meyer CF, Swenson BL, van Deursen JM, Baker DJ. Clearance of senescent glial cells prevents tau-dependent pathology and cognitive decline. Nature. 2018;562(7728):578–582. doi: 10.1038/s41586-018-0543-y.
-
DOI
-
PMC
-
PubMed
Childs BG, Baker DJ, Wijshake T, Conover CA, Campisi J, van Deursen JM. Senescent intimal foam cells are deleterious at all stages of atherosclerosis. Science. 2016;354(6311):472–477. doi: 10.1126/science.aaf6659.
-
DOI
-
PMC
-
PubMed
Demaria M, et al. An essential role for senescent cells in optimal wound healing through secretion of PDGF-AA. Dev Cell. 2014;31(6):722–733. doi: 10.1016/j.devcel.2014.11.012.
-
DOI
-
PMC
-
PubMed