

HEAT SHOCK FACTOR A8a Modulates Flavonoid Synthesis and Drought Tolerance
- PMID: 32958560
- PMCID: PMC7608180
- DOI: 10.1104/pp.20.01106
HEAT SHOCK FACTOR A8a Modulates Flavonoid Synthesis and Drought Tolerance
Abstract
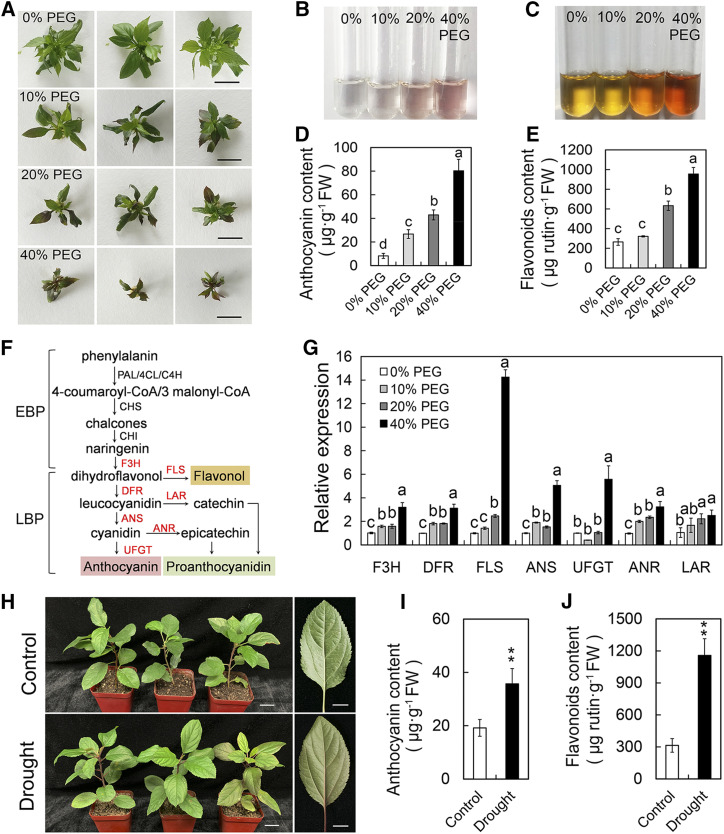
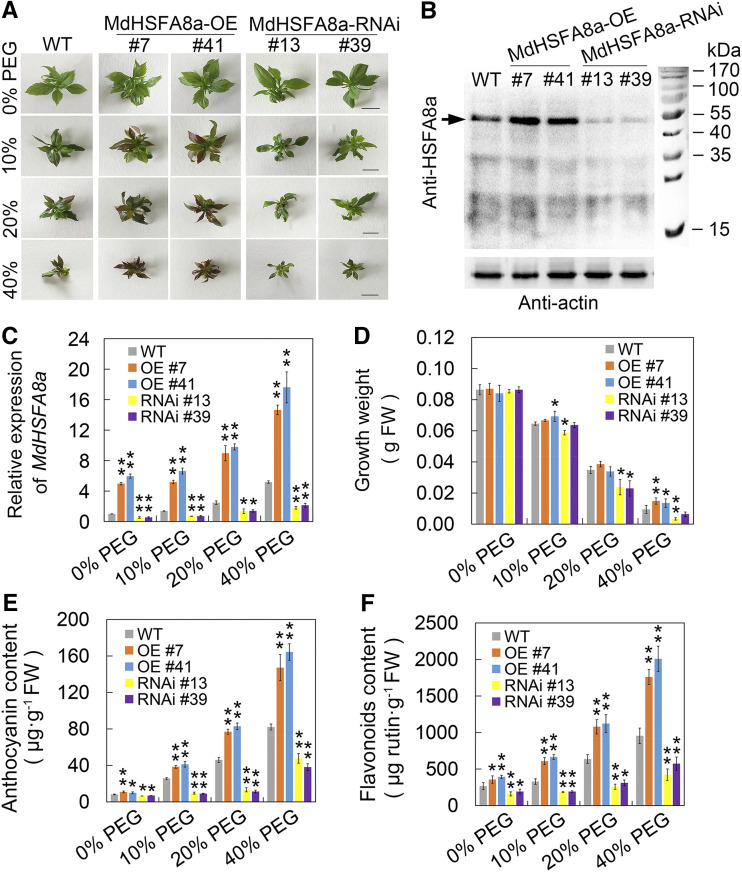
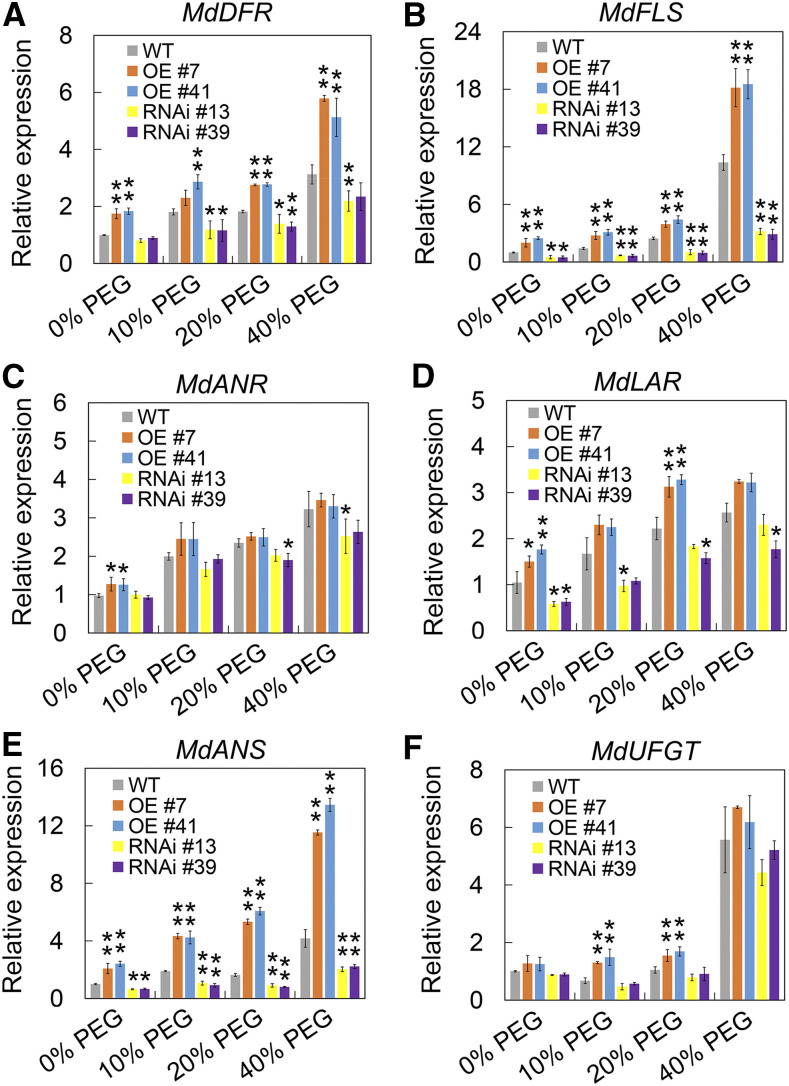
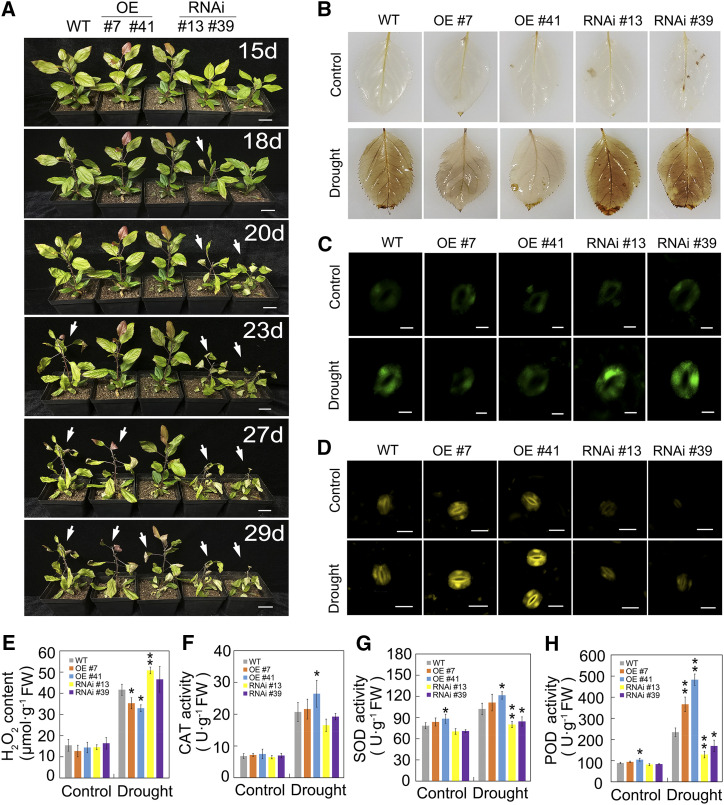
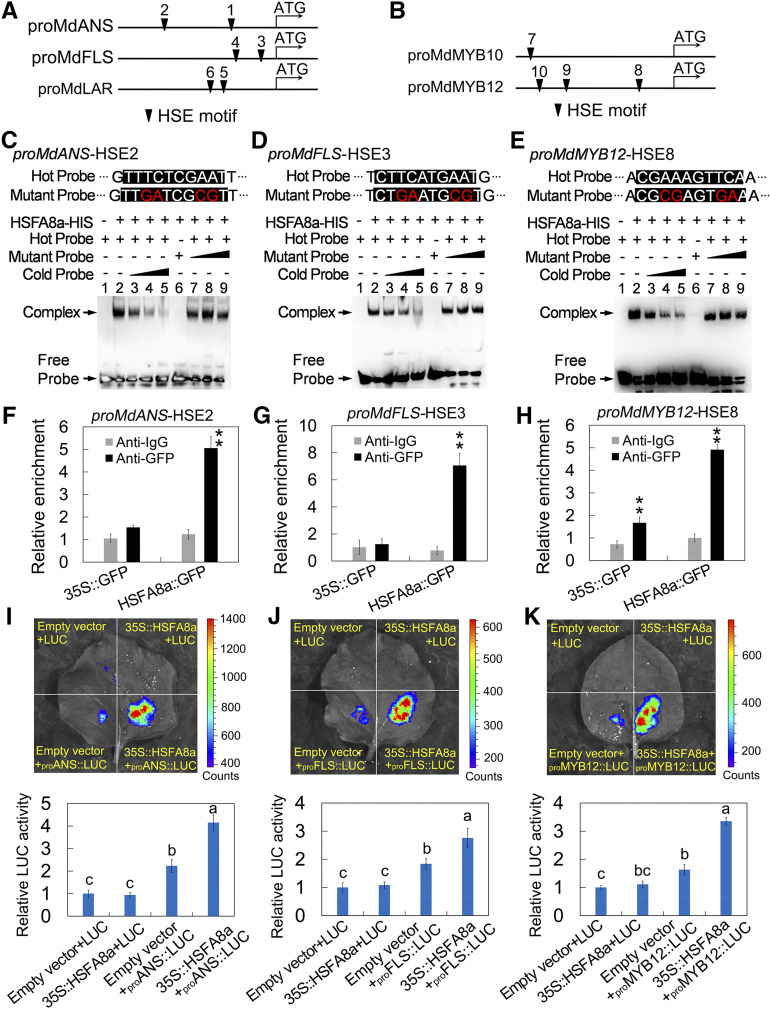
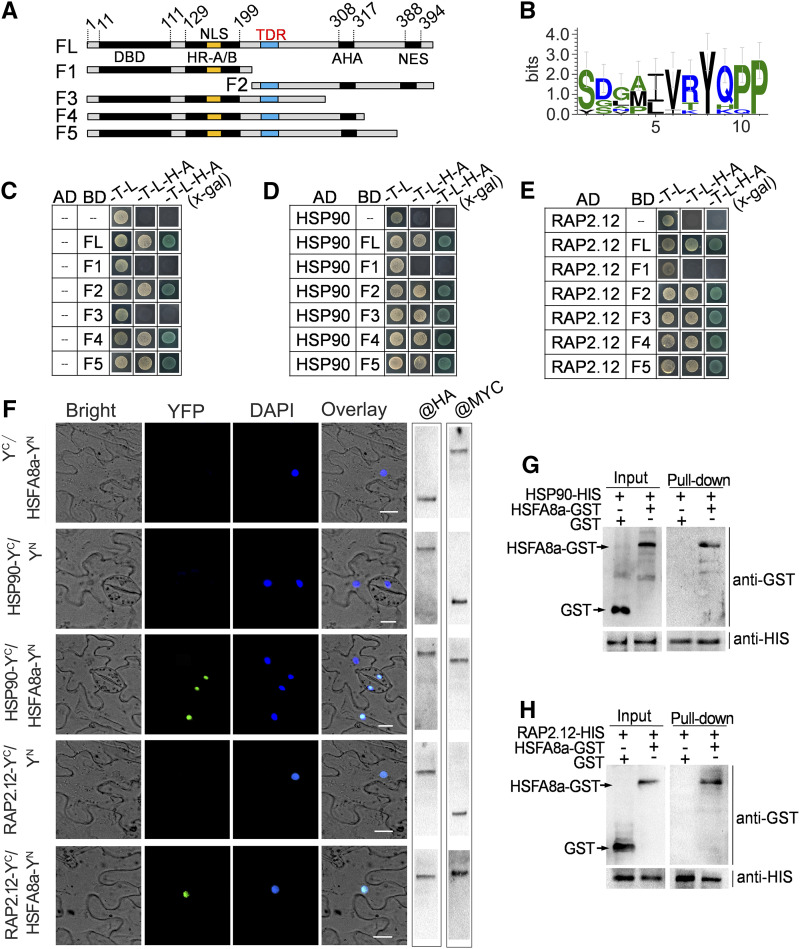
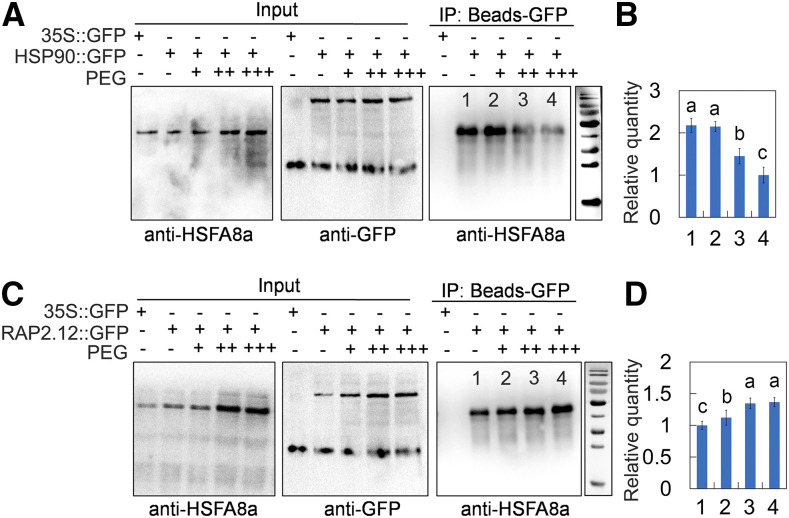
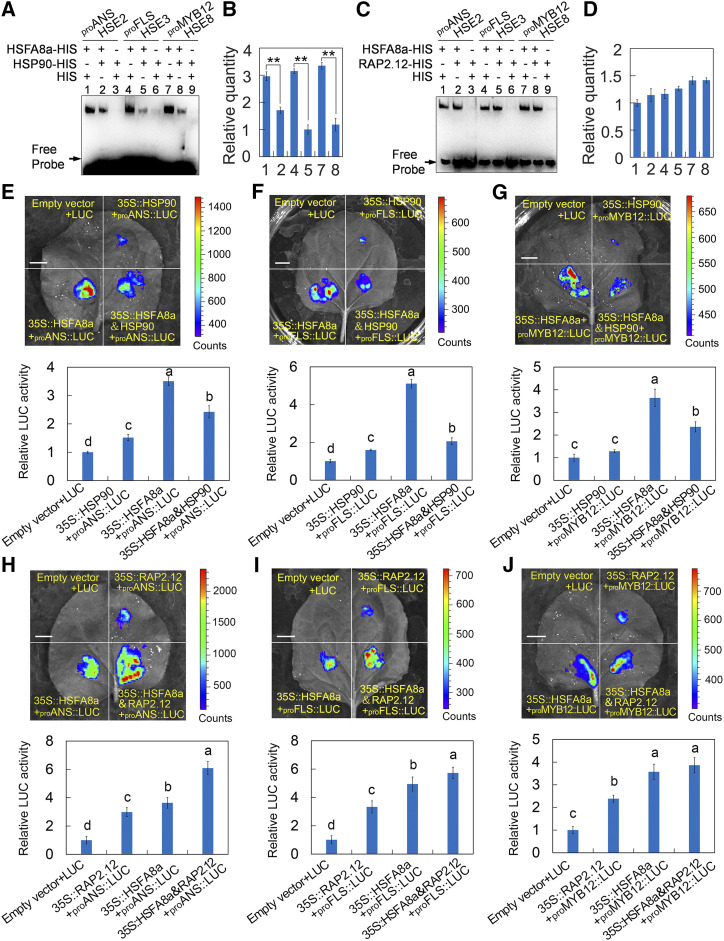
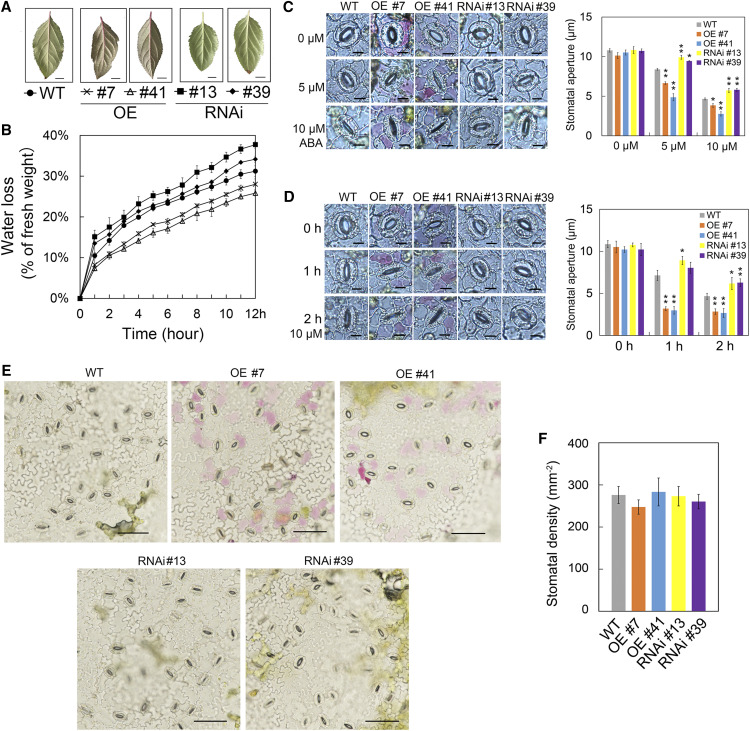
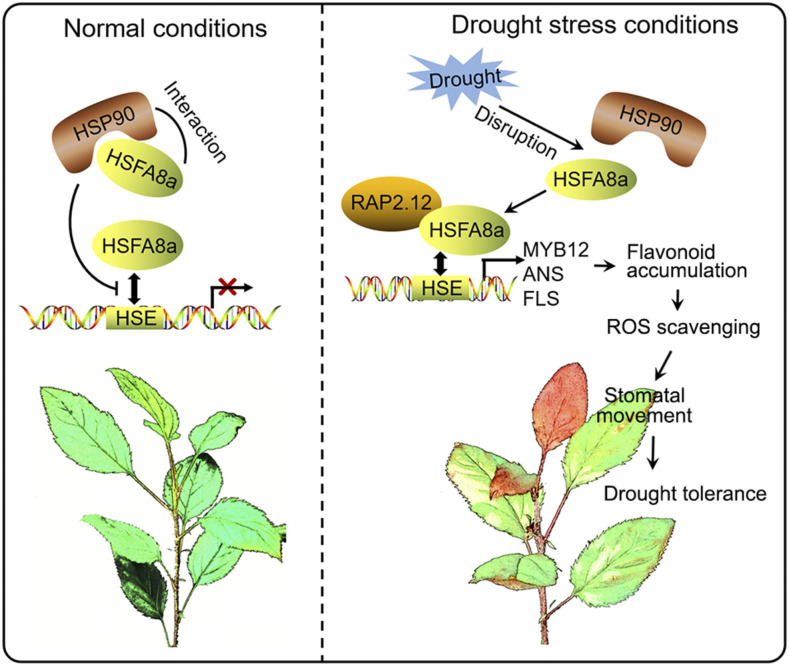
Drought is an important environmental factor affecting the growth and production of agricultural crops and fruits worldwide, including apple (Malus domestica). Heat shock factors (HSFs) have well-documented functions in stress responses, but their roles in flavonoid synthesis and the flavonoid-mediated drought response mechanism remain elusive. In this study, we demonstrated that a drought-responsive HSF, designated MdHSFA8a, promotes the accumulation of flavonoids, scavenging of reactive oxygen species, and plant survival under drought conditions. A chaperone, HEAT SHOCK PROTEIN90 (HSP90), interacted with MdHSFA8a to inhibit its binding activity and transcriptional activation. However, under drought stress, the MdHSP90-MdHSFA8a complex dissociated and the released MdHSFA8a further interacted with the APETALA2/ETHYLENE RESPONSIVE FACTOR family transcription factor RELATED TO AP2.12 to activate downstream gene activity. In addition, we demonstrated that MdHSFA8a participates in abscisic acid-induced stomatal closure and promotes the expression of abscisic acid signaling-related genes. Collectively, these findings provide insight into the mechanism by which stress-inducible MdHSFA8a modulates flavonoid synthesis to regulate drought tolerance.
© 2020 American Society of Plant Biologists. All Rights Reserved.
Figures
Similar articles
-
Heat-stress-responsive HvHSFA2e gene regulates the heat and drought tolerance in barley through modulation of phytohormone and secondary metabolic pathways.Plant Cell Rep. 2024 Jun 14;43(7):172. doi: 10.1007/s00299-024-03251-6. Plant Cell Rep. 2024. PMID: 38874775
-
Overexpression of MdATG8i Enhances Drought Tolerance by Alleviating Oxidative Damage and Promoting Water Uptake in Transgenic Apple.Int J Mol Sci. 2021 May 24;22(11):5517. doi: 10.3390/ijms22115517. Int J Mol Sci. 2021. PMID: 34073724 Free PMC article.
-
HISTONE DEACETYLASE 6 interaction with ABSCISIC ACID-INSENSITIVE 5 decreases apple drought tolerance.Plant Physiol. 2023 Nov 22;193(4):2711-2733. doi: 10.1093/plphys/kiad468. Plant Physiol. 2023. PMID: 37607253 Free PMC article.
-
Complex plant responses to drought and heat stress under climate change.Plant J. 2024 Mar;117(6):1873-1892. doi: 10.1111/tpj.16612. Epub 2024 Jan 3. Plant J. 2024. PMID: 38168757 Review.
-
Transcription Factors Interact with ABA through Gene Expression and Signaling Pathways to Mitigate Drought and Salinity Stress.Biomolecules. 2021 Aug 5;11(8):1159. doi: 10.3390/biom11081159. Biomolecules. 2021. PMID: 34439825 Free PMC article. Review.
Cited by
-
Functional Characterization of Heat Shock Factor (CrHsf) Families Provide Comprehensive Insight into the Adaptive Mechanisms of Canavalia rosea (Sw.) DC. to Tropical Coral Islands.Int J Mol Sci. 2022 Oct 15;23(20):12357. doi: 10.3390/ijms232012357. Int J Mol Sci. 2022. PMID: 36293211 Free PMC article.
-
MdNup62 involved in salt and osmotic stress tolerance in apple.Sci Rep. 2023 Nov 18;13(1):20198. doi: 10.1038/s41598-023-47024-9. Sci Rep. 2023. PMID: 37980385 Free PMC article.
-
PagMYB151 facilitates proline accumulation to enhance salt tolerance of poplar.BMC Genomics. 2023 Jun 22;24(1):345. doi: 10.1186/s12864-023-09459-2. BMC Genomics. 2023. PMID: 37349699 Free PMC article.
-
Phosphorylation of MdCYTOKININ RESPONSE FACTOR4 suppresses ethylene biosynthesis during apple fruit ripening.Plant Physiol. 2023 Jan 2;191(1):694-714. doi: 10.1093/plphys/kiac498. Plant Physiol. 2023. PMID: 36287070 Free PMC article.
-
A Functional InDel in the WRKY10 Promoter Controls the Degree of Flesh Red Pigmentation in Apple.Adv Sci (Weinh). 2024 Aug;11(30):e2400998. doi: 10.1002/advs.202400998. Epub 2024 Jun 14. Adv Sci (Weinh). 2024. PMID: 38874015 Free PMC article.
References
-
- Agati G, Tattini M(2010) Multiple functional roles of flavonoids in photoprotection. New Phytol 186: 786–793 - PubMed
-
- An JP, Li R, Qu FJ, You CX, Wang XF, Hao YJ(2018) R2R3-MYB transcription factor MdMYB23 is involved in the cold tolerance and proanthocyanidin accumulation in apple. Plant J 96: 562–577 - PubMed
-
- An XH, Tian Y, Chen KQ, Liu XJ, Liu DD, Xie XB, Cheng CG, Cong PH, Hao YJ(2015) MdMYB9 and MdMYB11 are involved in the regulation of the JA-induced biosynthesis of anthocyanin and proanthocyanidin in apples. Plant Cell Physiol 56: 650–662 - PubMed
-
- André CM, Schafleitner R, Legay S, Lefèvre I, Aliaga CAA, Nomberto G, Hoffmann L, Hausman JF, Larondelle Y, Evers D(2009) Gene expression changes related to the production of phenolic compounds in potato tubers grown under drought stress. Phytochemistry 70: 1107–1116 - PubMed
-
- Baniwal SK, Chan KY, Scharf KD, Nover L(2007) Role of heat stress transcription factor HsfA5 as specific repressor of HsfA4. J Biol Chem 282: 3605–3613 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
