

Quality-control mechanisms targeting translationally stalled and C-terminally extended poly(GR) associated with ALS/FTD
- PMID: 32958650
- PMCID: PMC7547246
- DOI: 10.1073/pnas.2005506117
Quality-control mechanisms targeting translationally stalled and C-terminally extended poly(GR) associated with ALS/FTD
Abstract
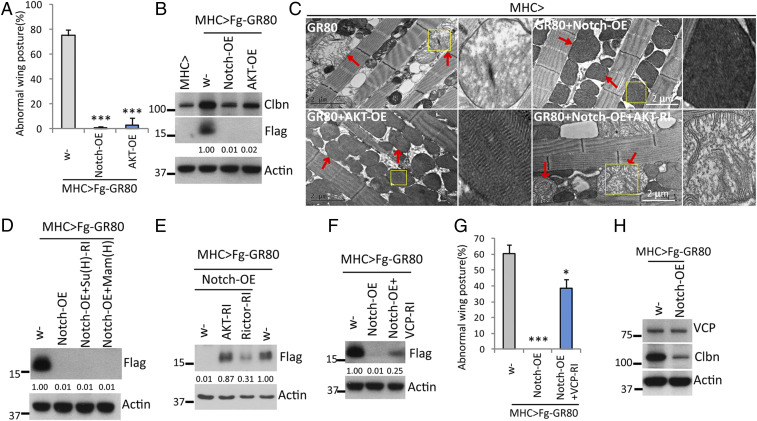
Maintaining the fidelity of nascent peptide chain (NP) synthesis is essential for proteome integrity and cellular health. Ribosome-associated quality control (RQC) serves to resolve stalled translation, during which untemplated Ala/Thr residues are added C terminally to stalled peptide, as shown during C-terminal Ala and Thr addition (CAT-tailing) in yeast. The mechanism and biological effects of CAT-tailing-like activity in metazoans remain unclear. Here we show that CAT-tailing-like modification of poly(GR), a dipeptide repeat derived from amyotrophic lateral sclerosis with frontotemporal dementia (ALS/FTD)-associated GGGGCC (G4C2) repeat expansion in C9ORF72, contributes to disease. We find that poly(GR) can act as a mitochondria-targeting signal, causing some poly(GR) to be cotranslationally imported into mitochondria. However, poly(GR) translation on mitochondrial surface is frequently stalled, triggering RQC and CAT-tailing-like C-terminal extension (CTE). CTE promotes poly(GR) stabilization, aggregation, and toxicity. Our genetic studies in Drosophila uncovered an important role of the mitochondrial protease YME1L in clearing poly(GR), revealing mitochondria as major sites of poly(GR) metabolism. Moreover, the mitochondria-associated noncanonical Notch signaling pathway impinges on the RQC machinery to restrain poly(GR) accumulation, at least in part through the AKT/VCP axis. The conserved actions of YME1L and noncanonical Notch signaling in animal models and patient cells support their fundamental involvement in ALS/FTD.
Keywords: C9-ALS/FTD; CAT-tailing; Notch; YME1L; ribosome-associated quality control.
Conflict of interest statement
The authors declare no competing interest.
Figures






Similar articles
-
ZNF598 co-translationally titrates poly(GR) protein implicated in the pathogenesis of C9ORF72-associated ALS/FTD.Nucleic Acids Res. 2021 Nov 8;49(19):11294-11311. doi: 10.1093/nar/gkab834. Nucleic Acids Res. 2021. PMID: 34551427 Free PMC article.
-
The mTORC2/AKT/VCP axis is associated with quality control of the stalled translation of poly(GR) dipeptide repeats in C9-ALS/FTD.J Biol Chem. 2023 Mar;299(3):102995. doi: 10.1016/j.jbc.2023.102995. Epub 2023 Feb 9. J Biol Chem. 2023. PMID: 36764521 Free PMC article.
-
Poly-GR repeats associated with ALS/FTD gene C9ORF72 impair translation elongation and induce a ribotoxic stress response in neurons.Sci Signal. 2024 Aug 6;17(848):eadl1030. doi: 10.1126/scisignal.adl1030. Epub 2024 Aug 6. Sci Signal. 2024. PMID: 39106320 Free PMC article.
-
Insights into C9ORF72-Related ALS/FTD from Drosophila and iPSC Models.Trends Neurosci. 2018 Jul;41(7):457-469. doi: 10.1016/j.tins.2018.04.002. Epub 2018 May 2. Trends Neurosci. 2018. PMID: 29729808 Free PMC article. Review.
-
All in the Family: Repeats and ALS/FTD.Trends Neurosci. 2018 May;41(5):247-250. doi: 10.1016/j.tins.2018.03.010. Trends Neurosci. 2018. PMID: 29703376 Free PMC article. Review.
Cited by
-
Mechanisms of Metal-Induced Mitochondrial Dysfunction in Neurological Disorders.Toxics. 2021 Jun 17;9(6):142. doi: 10.3390/toxics9060142. Toxics. 2021. PMID: 34204190 Free PMC article. Review.
-
Translational regulation by ribosome-associated quality control in neurodegenerative disease, cancer, and viral infection.Front Cell Dev Biol. 2022 Sep 14;10:970654. doi: 10.3389/fcell.2022.970654. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36187485 Free PMC article. Review.
-
Aloperine inhibits the progression of non-small-cell lung cancer through the PI3K/Akt signaling pathway.Cancer Cell Int. 2021 Dec 11;21(1):662. doi: 10.1186/s12935-021-02361-5. Cancer Cell Int. 2021. PMID: 34895234 Free PMC article.
-
Prevention of ribosome collision-induced neuromuscular degeneration by SARS CoV-2-encoded Nsp1.Proc Natl Acad Sci U S A. 2022 Oct 18;119(42):e2202322119. doi: 10.1073/pnas.2202322119. Epub 2022 Sep 28. Proc Natl Acad Sci U S A. 2022. PMID: 36170200 Free PMC article.
-
Inefficient quality control of ribosome stalling during APP synthesis generates CAT-tailed species that precipitate hallmarks of Alzheimer's disease.Acta Neuropathol Commun. 2021 Oct 18;9(1):169. doi: 10.1186/s40478-021-01268-6. Acta Neuropathol Commun. 2021. PMID: 34663454 Free PMC article.
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
