

Transcriptional Regulation of Wnt/β-Catenin Pathway in Colorectal Cancer
- PMID: 32961708
- PMCID: PMC7564852
- DOI: 10.3390/cells9092125
Transcriptional Regulation of Wnt/β-Catenin Pathway in Colorectal Cancer
Abstract
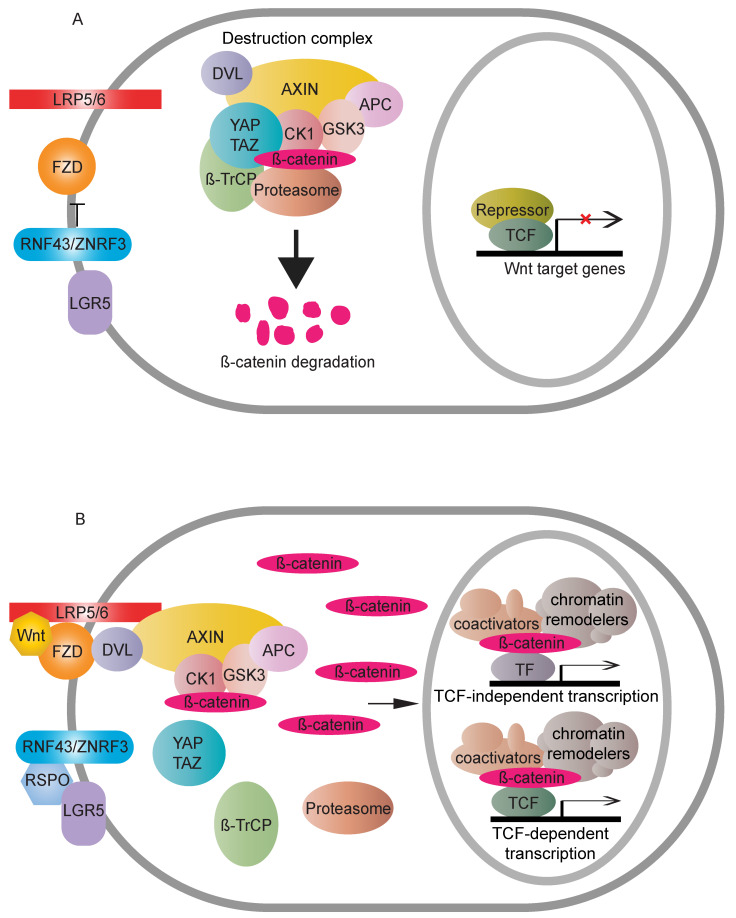
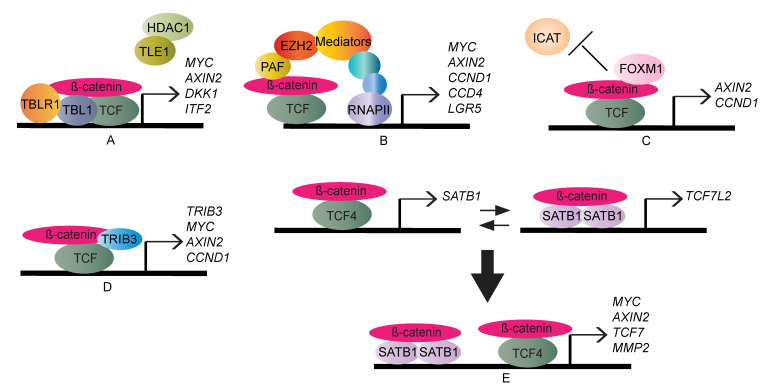
The Wnt/β-catenin signaling pathway exerts integral roles in embryogenesis and adult homeostasis. Aberrant activation of the pathway is implicated in growth-associated diseases and cancers, especially as a key driver in the initiation and progression of colorectal cancer (CRC). Loss or inactivation of Adenomatous polyposis coli (APC) results in constitutive activation of Wnt/β-catenin signaling, which is considered as an initiating event in the development of CRC. Increased Wnt/β-catenin signaling is observed in virtually all CRC patients, underscoring the importance of this pathway for therapeutic intervention. Prior studies have deciphered the regulatory networks required for the cytoplasmic stabilisation or degradation of the Wnt pathway effector, β-catenin. However, the mechanism whereby nuclear β-catenin drives or inhibits expression of Wnt target genes is more diverse and less well characterised. Here, we describe a brief synopsis of the core canonical Wnt pathway components, set the spotlight on nuclear mediators and highlight the emerging role of chromatin regulators as modulators of β-catenin-dependent transcription activity and oncogenic output.
Keywords: Wnt/β-catenin signaling pathway; colorectal cancer; epigenetic regulation; transcriptional regulation.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures



References
-
- Hart M., Concordet J.P., Lassot I., Albert I., del los Santos R., Durand H., Perret C., Rubinfeld B., Margottin F., Benarous R., et al. The F-box protein beta-TrCP associates with phosphorylated beta-catenin and regulates its activity in the cell. Curr. Biol. 1999;9:207–210. doi: 10.1016/S0960-9822(99)80091-8. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
