

Metabolism of Glycosphingolipids and Their Role in the Pathophysiology of Lysosomal Storage Disorders
- PMID: 32961778
- PMCID: PMC7555265
- DOI: 10.3390/ijms21186881
Metabolism of Glycosphingolipids and Their Role in the Pathophysiology of Lysosomal Storage Disorders
Abstract
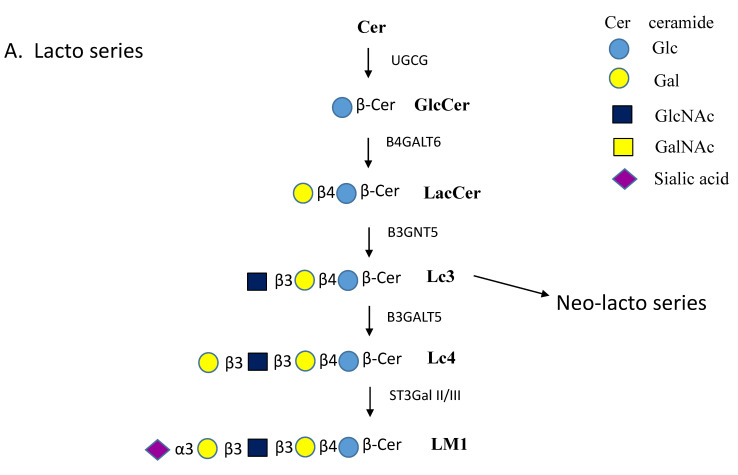
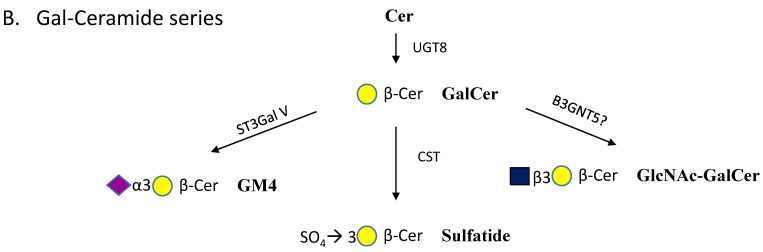
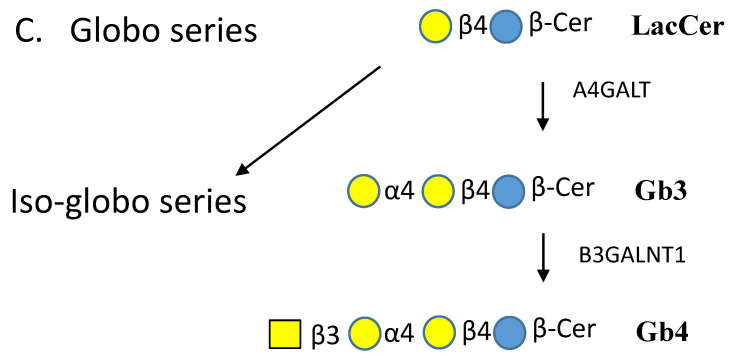
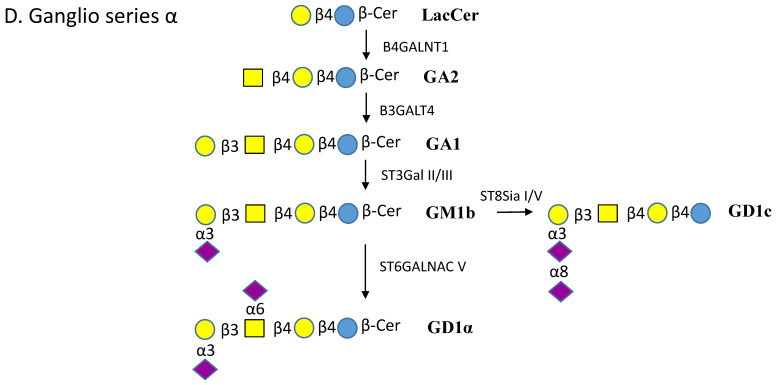
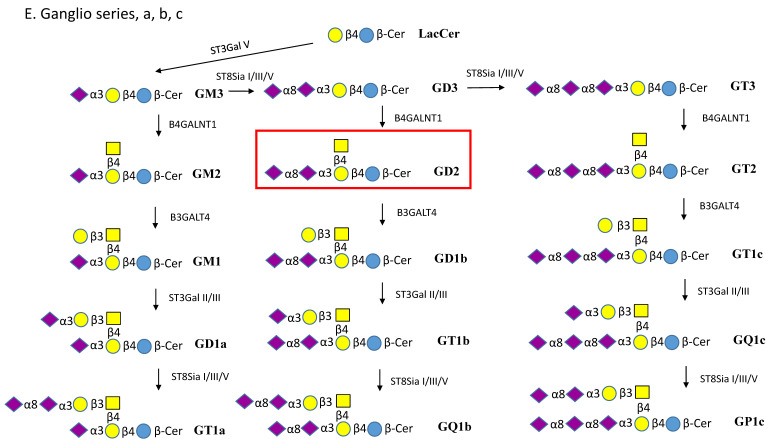
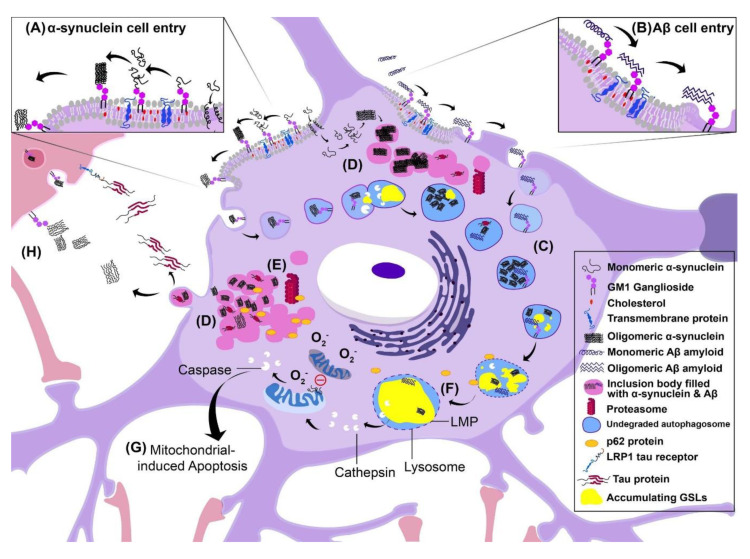
Glycosphingolipids (GSLs) are a specialized class of membrane lipids composed of a ceramide backbone and a carbohydrate-rich head group. GSLs populate lipid rafts of the cell membrane of eukaryotic cells, and serve important cellular functions including control of cell-cell signaling, signal transduction and cell recognition. Of the hundreds of unique GSL structures, anionic gangliosides are the most heavily implicated in the pathogenesis of lysosomal storage diseases (LSDs) such as Tay-Sachs and Sandhoff disease. Each LSD is characterized by the accumulation of GSLs in the lysosomes of neurons, which negatively interact with other intracellular molecules to culminate in cell death. In this review, we summarize the biosynthesis and degradation pathways of GSLs, discuss how aberrant GSL metabolism contributes to key features of LSD pathophysiology, draw parallels between LSDs and neurodegenerative proteinopathies such as Alzheimer's and Parkinson's disease and lastly, discuss possible therapies for patients.
Keywords: biosynthesis; degradation of glycoconjugates; glycosphingolipids; glycosyl hydrolases; lysosomal storage diseases; pathophysiology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
