

Minimal Elements Required for the Formation of Respiratory Syncytial Virus Cytoplasmic Inclusion Bodies In Vivo and In Vitro
- PMID: 32963000
- PMCID: PMC7512546
- DOI: 10.1128/mBio.01202-20
Minimal Elements Required for the Formation of Respiratory Syncytial Virus Cytoplasmic Inclusion Bodies In Vivo and In Vitro
Abstract
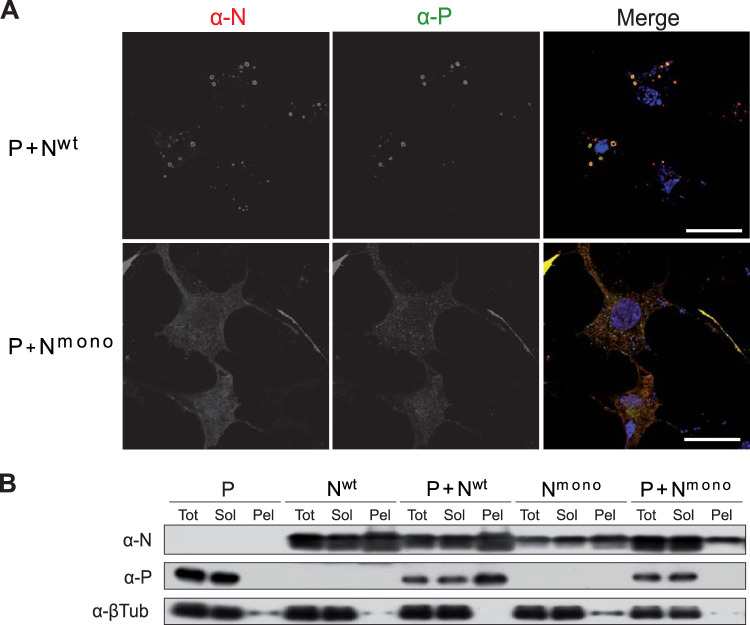
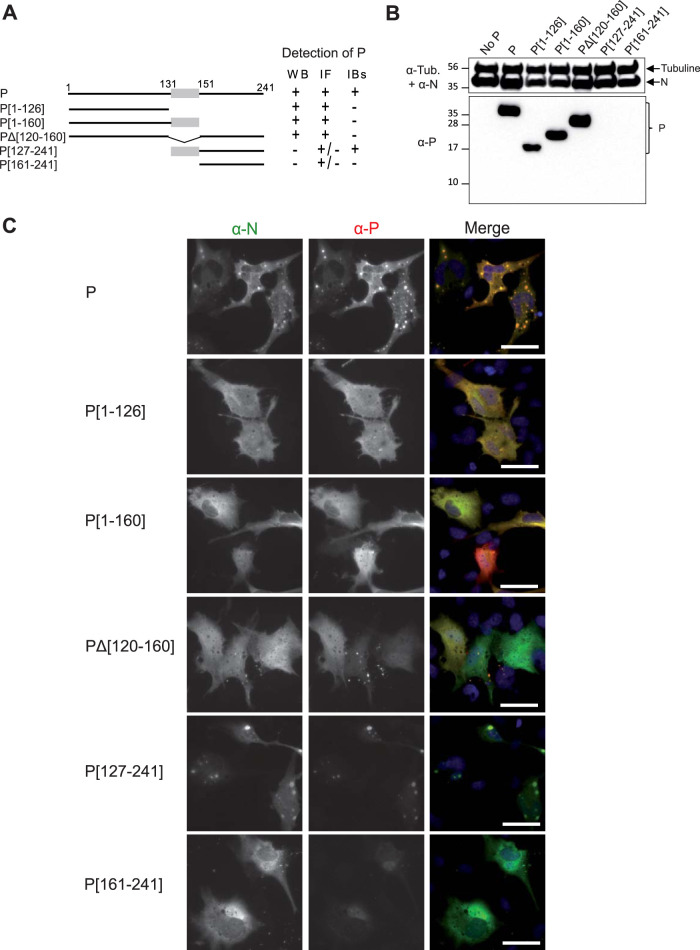
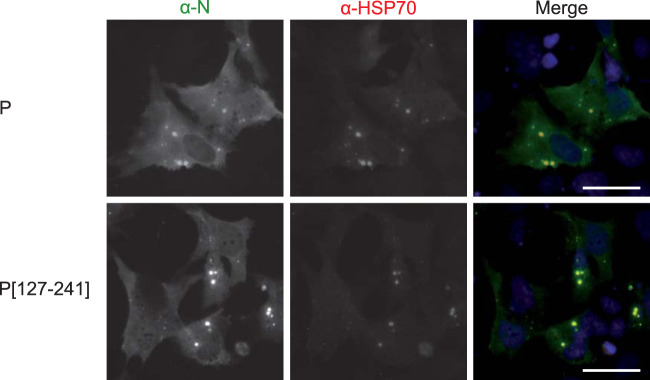
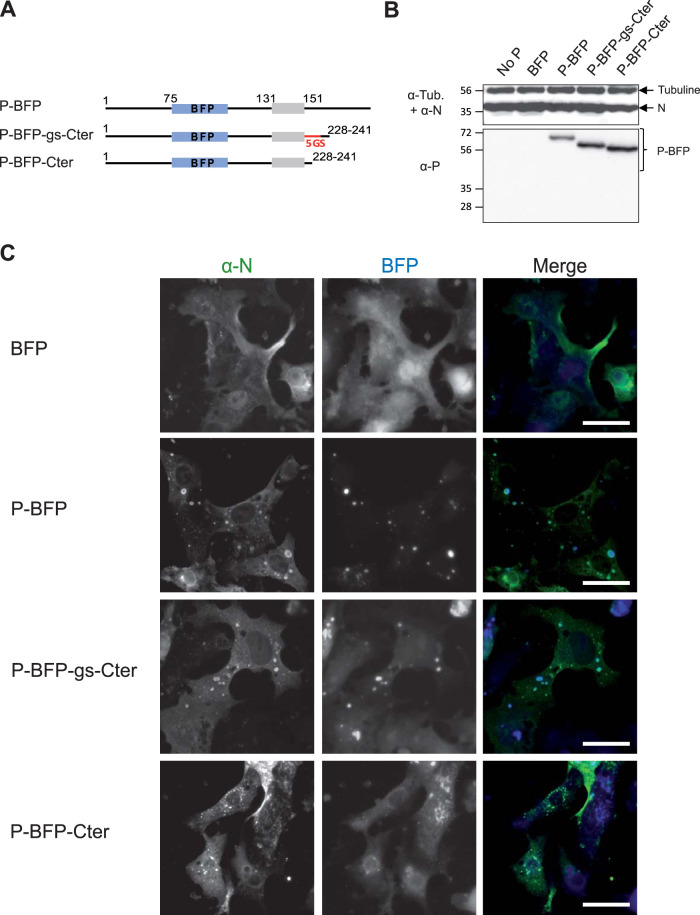
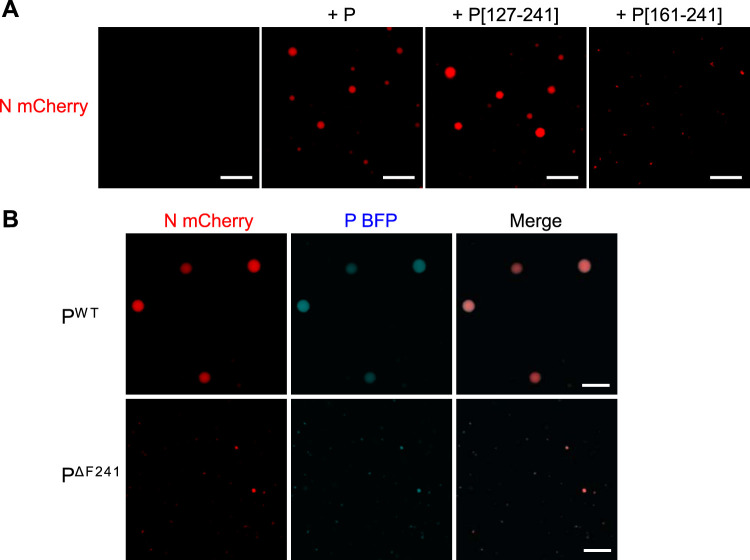
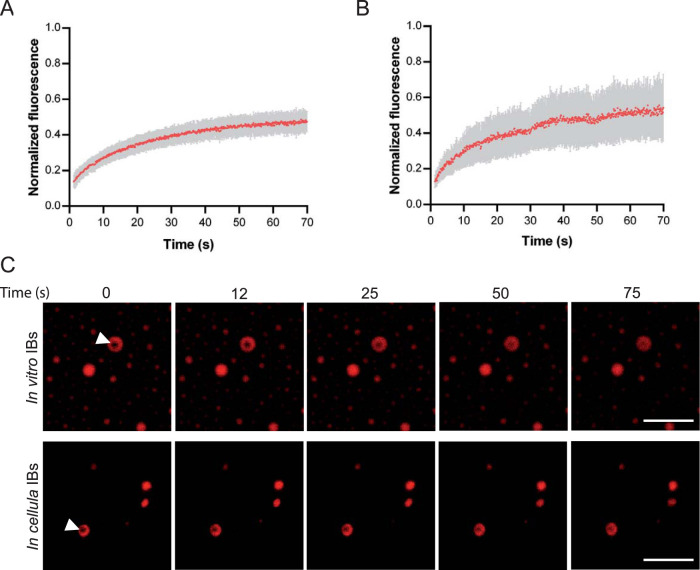
Infection of host cells by the respiratory syncytial virus (RSV) is characterized by the formation of spherical cytoplasmic inclusion bodies (IBs). These structures, which concentrate all the proteins of the polymerase complex as well as some cellular proteins, were initially considered aggresomes formed by viral dead-end products. However, recent studies revealed that IBs are viral factories where viral RNA synthesis, i.e., replication and transcription, occurs. The analysis of IBs by electron microscopy revealed that they are membrane-less structures, and accumulated data on their structure, organization, and kinetics of formation revealed that IBs share the characteristics of cellular organelles, such as P-bodies or stress granules, suggesting that their morphogenesis depends on a liquid-liquid phase separation mechanism. It was previously shown that expression of the RSV nucleoprotein N and phosphoprotein P of the polymerase complex is sufficient to induce the formation of pseudo-IBs. Here, using a series of truncated P proteins, we identified the domains of P required for IB formation and show that the oligomeric state of N, provided it can interact with RNA, is critical for their morphogenesis. We also show that pseudo-IBs can form in vitro when recombinant N and P proteins are mixed. Finally, using fluorescence recovery after photobleaching approaches, we reveal that in cellula and in vitro IBs are liquid organelles. Our results strongly support the liquid-liquid phase separation nature of IBs and pave the way for further characterization of their dynamics.IMPORTANCE Respiratory syncytial virus (RSV) is the leading cause of lower respiratory tract illness in infants, elderly, and immunocompromised people. No vaccine or efficient antiviral treatment is available against this virus. The replication and transcription steps of the viral genome are appealing mechanisms to target for the development of new antiviral strategies. These activities take place within cytoplasmic inclusion bodies (IBs) that assemble during infection. Although expression of both the viral nucleoprotein (N) and phosphoprotein (P) allows induction of the formation of these IBs, the mechanism sustaining their assembly remains poorly characterized. Here, we identified key elements of N and P required for the scaffolding of IBs and managed for the first time to reconstitute RSV pseudo-IBs in vitro by coincubating recombinant N and P proteins. Our results provide strong evidence that the biogenesis of RSV IBs occurs through liquid-liquid phase transition mediated by N-P interactions.
Keywords: RSV; inclusion bodies; liquid-liquid phase; nucleoprotein; phosphoprotein; protein-protein interactions.
Copyright © 2020 Galloux et al.
Figures

References
-
- Afonso CL, Amarasinghe GK, Bányai K, Bào Y, Basler CF, Bavari S, Bejerman N, Blasdell KR, Briand F-X, Briese T, Bukreyev A, Calisher CH, Chandran K, Chéng J, Clawson AN, Collins PL, Dietzgen RG, Dolnik O, Domier LL, Dürrwald R, Dye JM, Easton AJ, Ebihara H, Farkas SL, Freitas-Astúa J, Formenty P, Fouchier RAM, Fù Y, Ghedin E, Goodin MM, Hewson R, Horie M, Hyndman TH, Jiāng D, Kitajima EW, Kobinger GP, Kondo H, Kurath G, Lamb RA, Lenardon S, Leroy EM, Li C-X, Lin X-D, Liú L, Longdon B, Marton S, Maisner A, Mühlberger E, Netesov SV, Nowotny N, et al. 2016. Taxonomy of the order Mononegavirales: update 2016. Arch Virol 161:2351–2360. doi:10.1007/s00705-016-2880-1. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
