

IL-33-ST2 axis regulates myeloid cell differentiation and activation enabling effective club cell regeneration
- PMID: 32963227
- PMCID: PMC7508874
- DOI: 10.1038/s41467-020-18466-w
IL-33-ST2 axis regulates myeloid cell differentiation and activation enabling effective club cell regeneration
Abstract
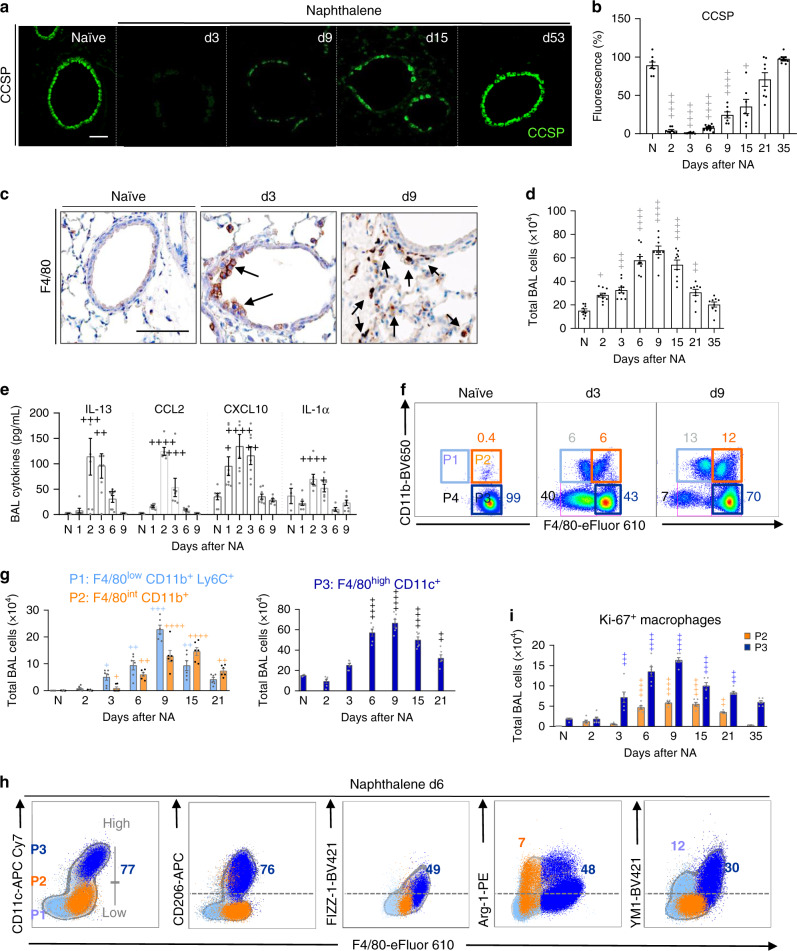
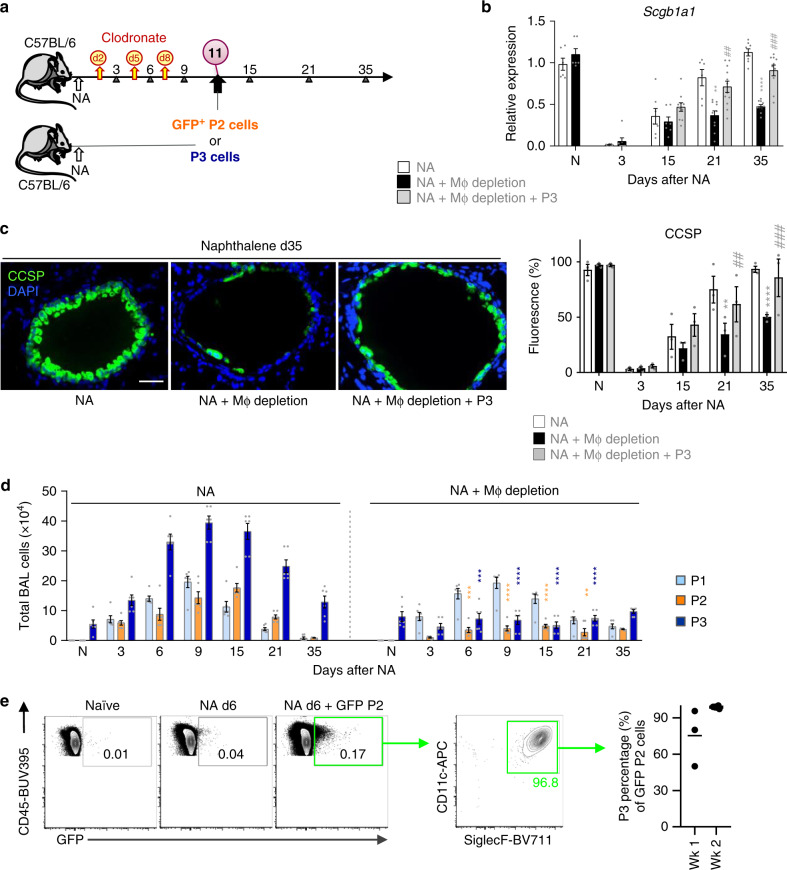
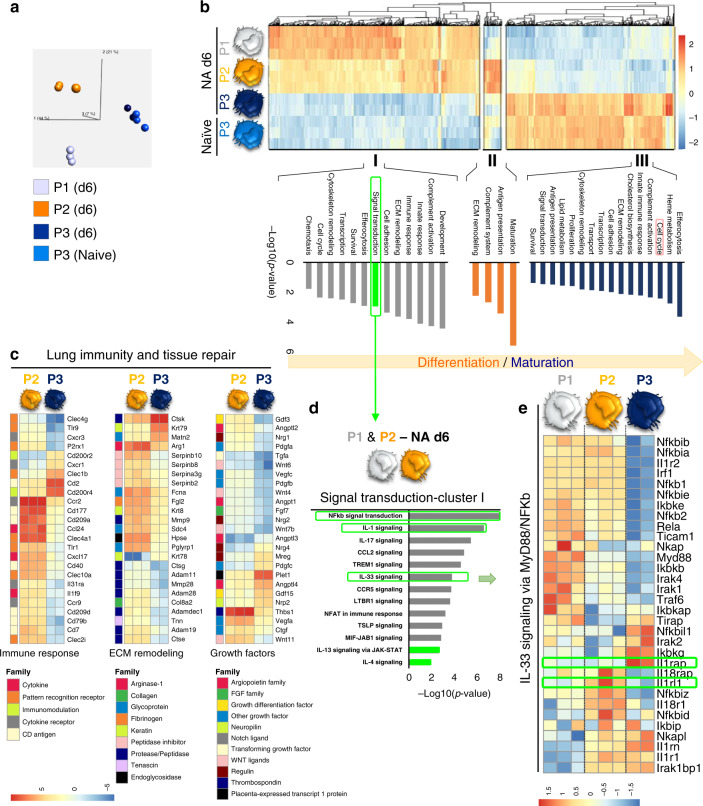
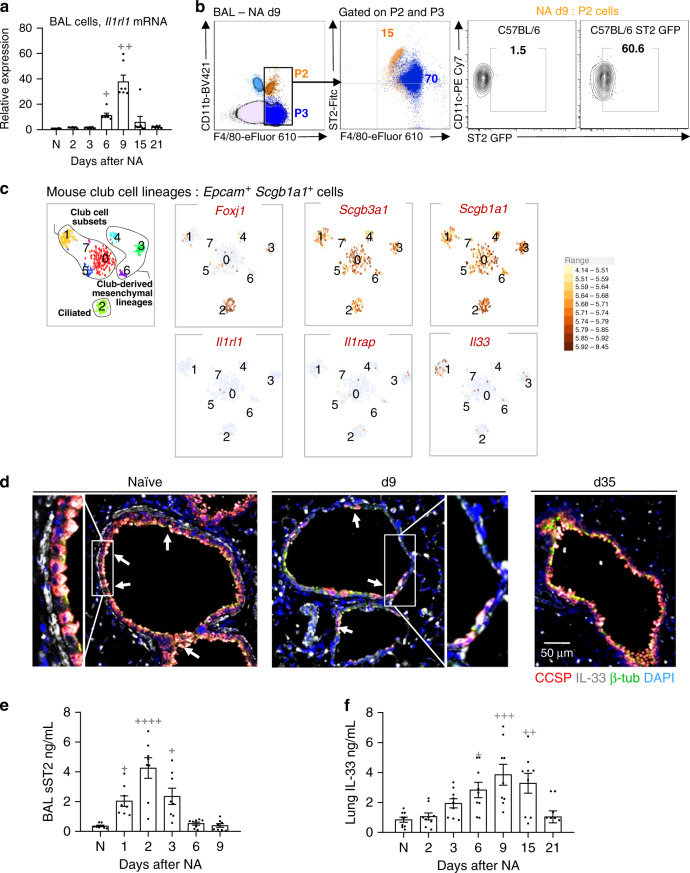
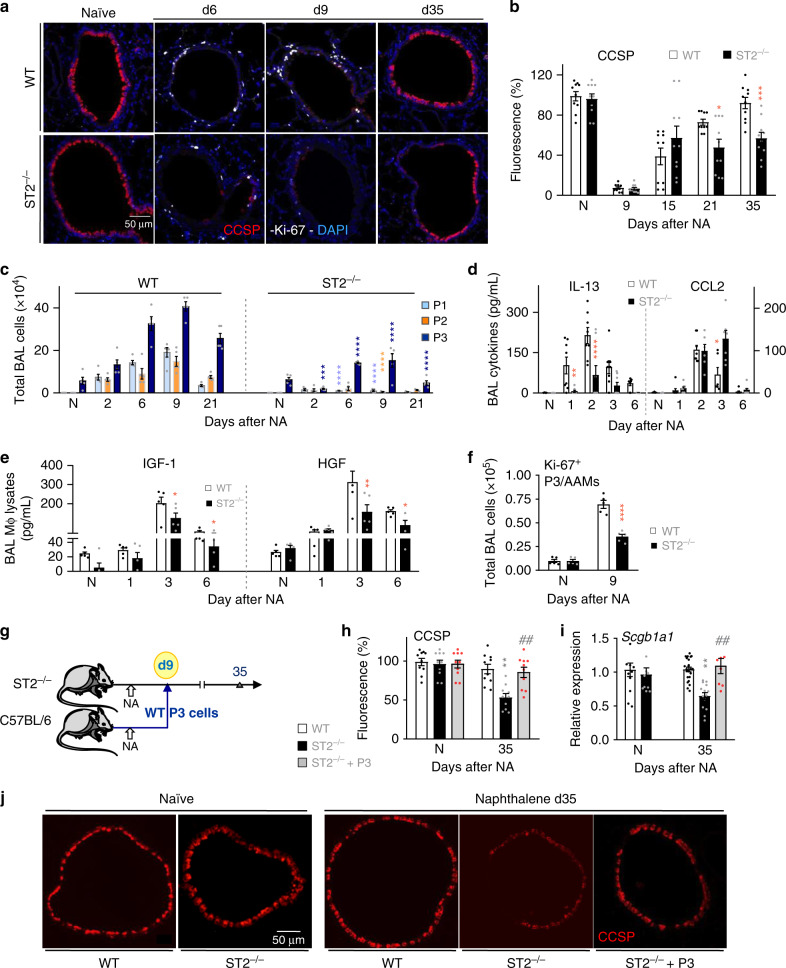
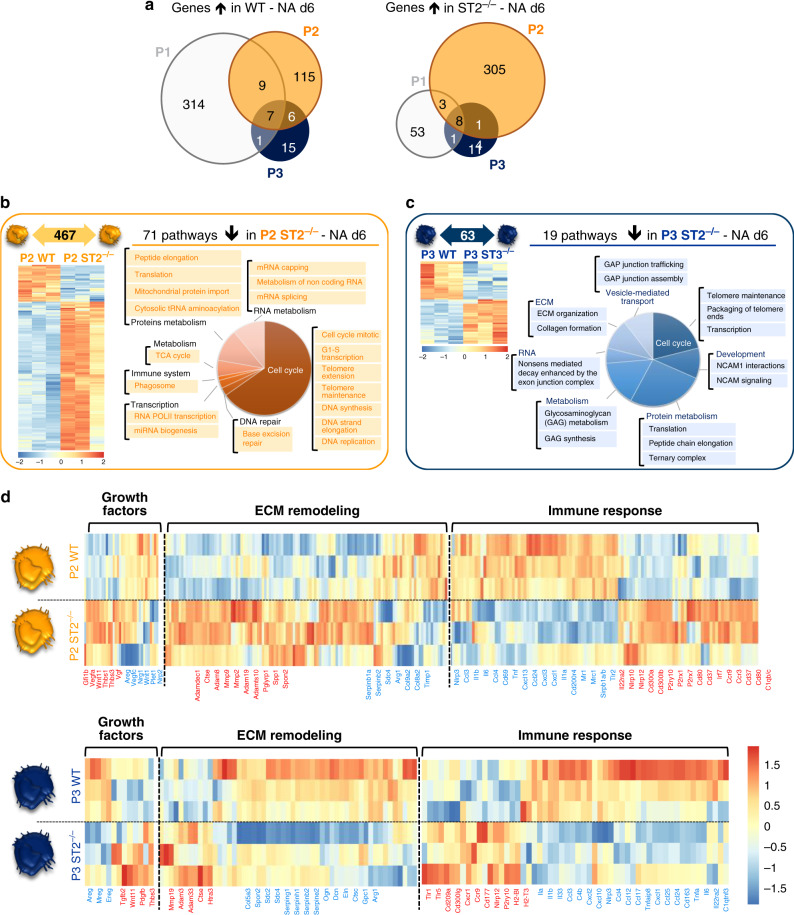
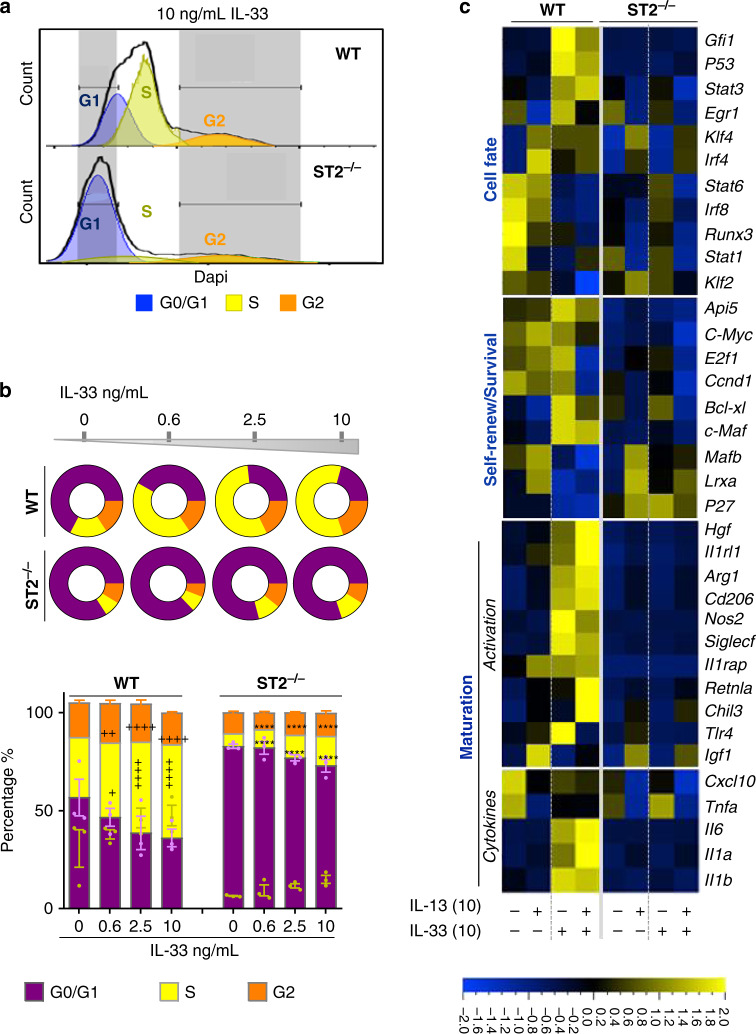
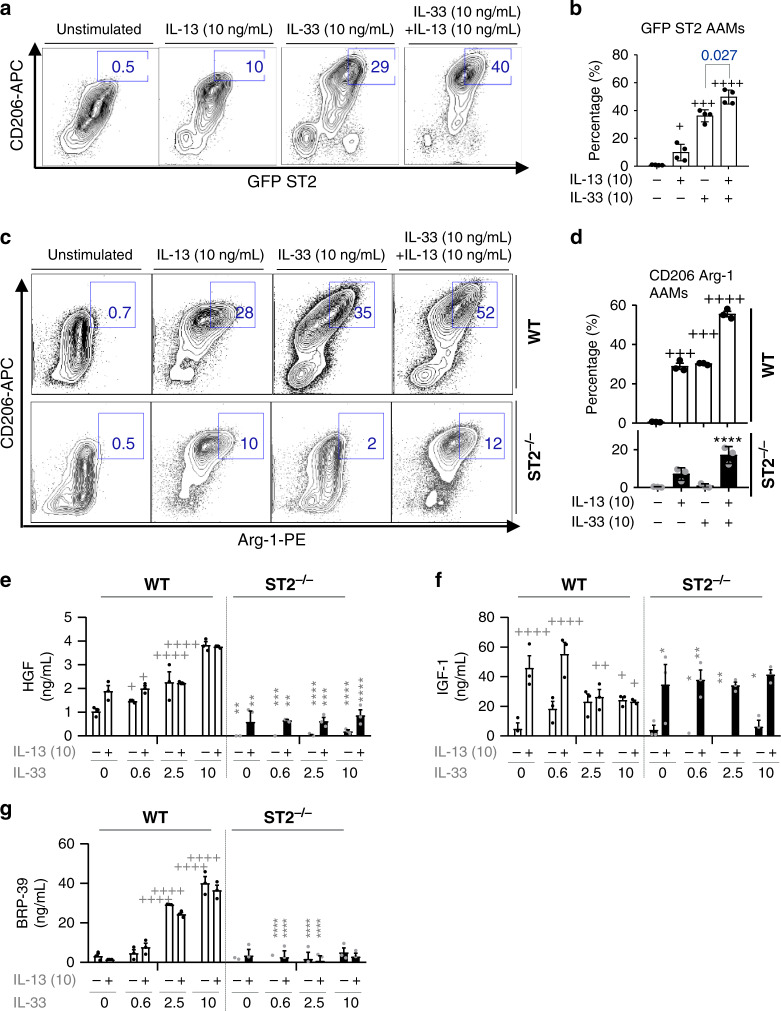
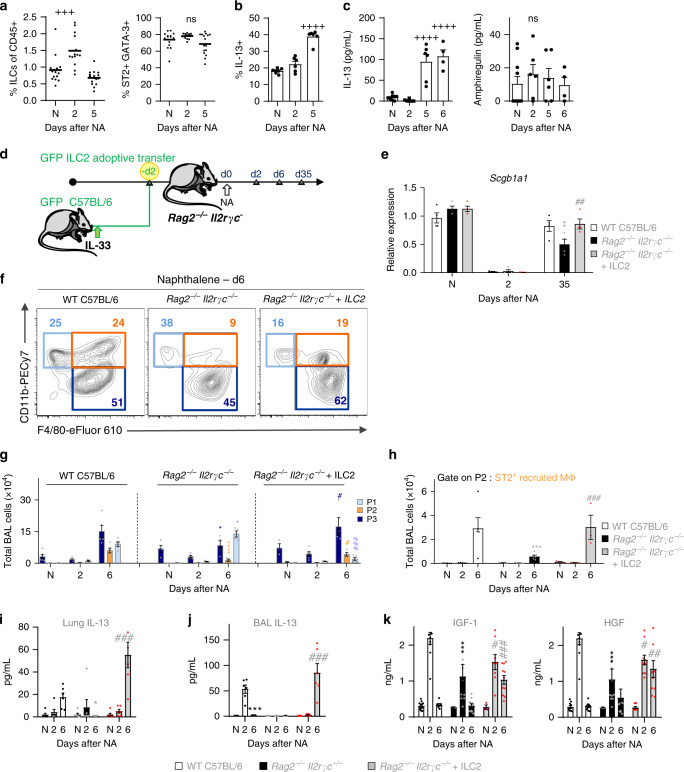
Evidence points to an indispensable function of macrophages in tissue regeneration, yet the underlying molecular mechanisms remain elusive. Here we demonstrate a protective function for the IL-33-ST2 axis in bronchial epithelial repair, and implicate ST2 in myeloid cell differentiation. ST2 deficiency in mice leads to reduced lung myeloid cell infiltration, abnormal alternatively activated macrophage (AAM) function, and impaired epithelial repair post naphthalene-induced injury. Reconstitution of wild type (WT) AAMs to ST2-deficient mice completely restores bronchial re-epithelialization. Central to this mechanism is the direct effect of IL-33-ST2 signaling on monocyte/macrophage differentiation, self-renewal and repairing ability, as evidenced by the downregulation of key pathways regulating myeloid cell cycle, maturation and regenerative function of the epithelial niche in ST2-/- mice. Thus, the IL-33-ST2 axis controls epithelial niche regeneration by activating a large multi-cellular circuit, including monocyte differentiation into competent repairing AAMs, as well as group-2 innate lymphoid cell (ILC2)-mediated AAM activation.
Conflict of interest statement
The authors R.D., A.M.C., A.B., J.W., J.W., X.Q., Y.S., R.R., R.K., and A.A.H. are/were employed by and shareholders of AstraZeneca. The author M.P. received funding from MedImmune/AstraZeneca. The remaining authors declare no competing interests.
Figures
Similar articles
-
An ST2-dependent role of bone marrow-derived group 2 innate lymphoid cells in pulmonary fibrosis.J Pathol. 2018 Aug;245(4):399-409. doi: 10.1002/path.5092. Epub 2018 Jun 5. J Pathol. 2018. PMID: 29722022
-
The interleukin-33 receptor ST2 is important for the development of peripheral airway hyperresponsiveness and inflammation in a house dust mite mouse model of asthma.Clin Exp Allergy. 2016 Mar;46(3):479-90. doi: 10.1111/cea.12683. Clin Exp Allergy. 2016. PMID: 26609909
-
IL-33/ST2 Axis in Organ Fibrosis.Front Immunol. 2018 Oct 24;9:2432. doi: 10.3389/fimmu.2018.02432. eCollection 2018. Front Immunol. 2018. PMID: 30405626 Free PMC article. Review.
-
Deficiency in IL-33/ST2 Axis Reshapes Mitochondrial Metabolism in Lipopolysaccharide-Stimulated Macrophages.Front Immunol. 2019 Feb 1;10:127. doi: 10.3389/fimmu.2019.00127. eCollection 2019. Front Immunol. 2019. PMID: 30774633 Free PMC article.
-
Regulatory Mechanisms of IL-33-ST2-Mediated Allergic Inflammation.Front Immunol. 2018 Sep 4;9:2004. doi: 10.3389/fimmu.2018.02004. eCollection 2018. Front Immunol. 2018. PMID: 30233590 Free PMC article. Review.
Cited by
-
IL1RL1 polymorphisms rs12479210 and rs1420101 are associated with increased lung cancer risk in the Chinese Han population.Front Genet. 2023 Aug 31;14:1183528. doi: 10.3389/fgene.2023.1183528. eCollection 2023. Front Genet. 2023. PMID: 37719702 Free PMC article.
-
LKB1 deficiency upregulates RELM-α to drive airway goblet cell metaplasia.Cell Mol Life Sci. 2021 Dec 18;79(1):42. doi: 10.1007/s00018-021-04044-w. Cell Mol Life Sci. 2021. PMID: 34921639 Free PMC article.
-
Expansion of ST2-expressing macrophages in a patient with bronchiolitis obliterans syndrome.ERJ Open Res. 2023 May 30;9(3):00033-2023. doi: 10.1183/23120541.00033-2023. eCollection 2023 May. ERJ Open Res. 2023. PMID: 37260458 Free PMC article.
-
M2 Macrophages promote IL-33 expression, ILC2 expansion and mucous metaplasia in response to early life rhinovirus infections.Front Immunol. 2022 Aug 12;13:952509. doi: 10.3389/fimmu.2022.952509. eCollection 2022. Front Immunol. 2022. PMID: 36032072 Free PMC article.
-
Maintenance of Barrier Tissue Integrity by Unconventional Lymphocytes.Front Immunol. 2021 Apr 14;12:670471. doi: 10.3389/fimmu.2021.670471. eCollection 2021. Front Immunol. 2021. PMID: 33936115 Free PMC article. Review.
References
-
- Hong KU, Reynolds SD, Giangreco A, Hurley CM, Stripp BR. Clara cell secretory protein-expressing cells of the airway neuroepithelial body microenvironment include a label-retaining subset and are critical for epithelial renewal after progenitor cell depletion. Am. J. Respir. Cell Mol. Biol. 2001;24:671–681. doi: 10.1165/ajrcmb.24.6.4498. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
