

Cannabidiol activation of vagal afferent neurons requires TRPA1
- PMID: 32965166
- PMCID: PMC8356782
- DOI: 10.1152/jn.00128.2020
Cannabidiol activation of vagal afferent neurons requires TRPA1
Abstract
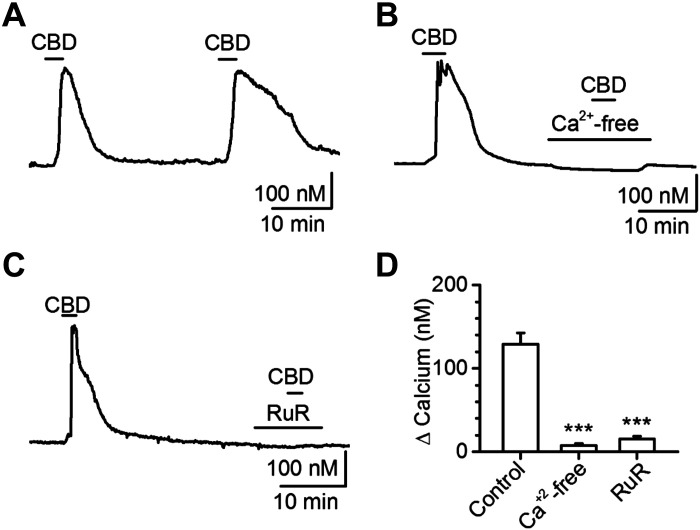
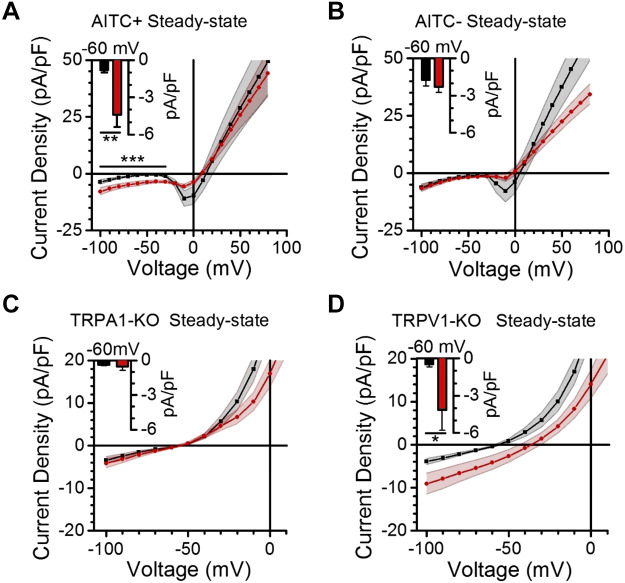
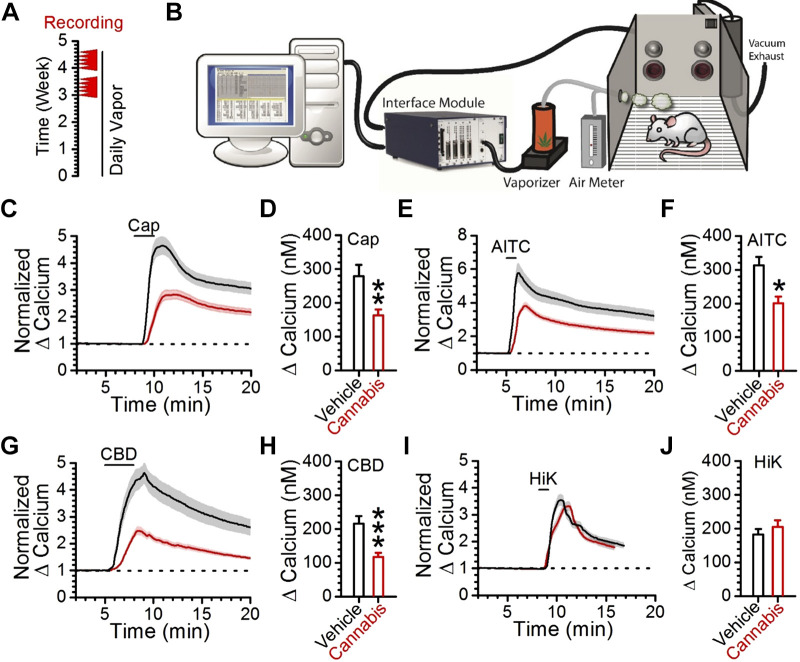
Vagal afferent neurons abundantly express excitatory transient receptor potential (TRP) channels, which strongly influence afferent signaling. Cannabinoids have been identified as direct agonists of TRP channels, including TRPA1 and TRPV1, suggesting that exogenous cannabinoids may influence vagal signaling via TRP channel activation. The diverse therapeutic effects of electrical vagus nerve stimulation also result from administration of the nonpsychotropic cannabinoid, cannabidiol (CBD); however, the direct effects of CBD on vagal afferent signaling remain unknown. We investigated actions of CBD on vagal afferent neurons, using calcium imaging and electrophysiology. CBD produced strong excitatory effects in neurons expressing TRPA1. CBD responses were prevented by removal of bath calcium, ruthenium red, and the TRPA1 antagonist A967079, but not the TRPV1 antagonist SB366791, suggesting an essential role for TRPA1. These pharmacological experiments were confirmed using genetic knockouts where TRPA1 KO mice lacked CBD responses, whereas TRPV1 knockout (KO) mice exhibited CBD-induced activation. We also characterized CBD-provoked inward currents at resting potentials in vagal afferents expressing TRPA1 that were absent in TRPA1 KO mice, but persisted in TRPV1 KO mice. CBD also inhibited voltage-activated sodium conductances in A-fiber, but not in C-fiber afferents. To simulate adaptation, resulting from chronic cannabis use, we administered cannabis extract vapor daily for 3 wk. Cannabis exposure reduced the magnitude of CBD responses, likely due to a loss of TRPA1 signaling. Together, these findings detail a novel excitatory action of CBD at vagal afferent neurons, which requires TRPA1 and may contribute to the vagal mimetic effects of CBD and adaptation following chronic cannabis use.NEW & NOTEWORTHY CBD usage has increased with its legalization. The clinical efficacy of CBD has been demonstrated for conditions including some forms of epilepsy, depression, and anxiety that are also treatable by vagus nerve stimulation. We found CBD exhibited direct excitatory effects on vagal afferent neurons that required TRPA1, were augmented by TRPV1, and attenuated following chronic cannabis vapor exposure. These effects may contribute to vagal mimetic effects of CBD and adaptation after chronic cannabis use.
Keywords: CBD; cannabis; nodose ganglia; transient receptor potential; vagus.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures





Similar articles
-
TRPA1 channels mediate cold temperature sensing in mammalian vagal sensory neurons: pharmacological and genetic evidence.J Neurosci. 2008 Jul 30;28(31):7863-75. doi: 10.1523/JNEUROSCI.1696-08.2008. J Neurosci. 2008. PMID: 18667618 Free PMC article.
-
Effect of high-fat diet on mechanosensitive transient receptor potential channel activation in vagal afferent neurons.Can J Physiol Pharmacol. 2021 Jun;99(6):660-666. doi: 10.1139/cjpp-2020-0306. Epub 2020 Oct 27. Can J Physiol Pharmacol. 2021. PMID: 33108741
-
Relative contributions of TRPA1 and TRPV1 channels in the activation of vagal bronchopulmonary C-fibres by the endogenous autacoid 4-oxononenal.J Physiol. 2008 Jul 15;586(14):3447-59. doi: 10.1113/jphysiol.2008.153585. Epub 2008 May 22. J Physiol. 2008. PMID: 18499726 Free PMC article.
-
Interaction between TRPA1 and TRPV1: Synergy on pulmonary sensory nerves.Pulm Pharmacol Ther. 2015 Dec;35:87-93. doi: 10.1016/j.pupt.2015.08.003. Epub 2015 Aug 14. Pulm Pharmacol Ther. 2015. PMID: 26283426 Free PMC article. Review.
-
Pharmacological effects of cannabidiol by transient receptor potential channels.Life Sci. 2022 Jul 1;300:120582. doi: 10.1016/j.lfs.2022.120582. Epub 2022 Apr 26. Life Sci. 2022. PMID: 35483477 Review.
Cited by
-
Regulation of transient receptor potential channels by traditional Chinese medicines and their active ingredients.Front Pharmacol. 2022 Oct 13;13:1039412. doi: 10.3389/fphar.2022.1039412. eCollection 2022. Front Pharmacol. 2022. PMID: 36313301 Free PMC article. Review.
-
CCK-sensitive C fibers activate NTS leptin receptor-expressing neurons via NMDA receptors.Am J Physiol Regul Integr Comp Physiol. 2024 May 1;326(5):R383-R400. doi: 10.1152/ajpregu.00238.2022. Epub 2023 Dec 18. Am J Physiol Regul Integr Comp Physiol. 2024. PMID: 38105761 Free PMC article.
-
Utilization of Cannabidiol in Post-Organ-Transplant Care.Int J Mol Sci. 2025 Jan 15;26(2):699. doi: 10.3390/ijms26020699. Int J Mol Sci. 2025. PMID: 39859413 Free PMC article. Review.
-
Transient Receptor Potential (TRP) and Thermoregulation in Animals: Structural Biology and Neurophysiological Aspects.Animals (Basel). 2022 Jan 2;12(1):106. doi: 10.3390/ani12010106. Animals (Basel). 2022. PMID: 35011212 Free PMC article. Review.
-
Role of Vagus Nerve Stimulation in Refractory and Super Refractory Status Epilepticus: A Pediatric Case Series.Brain Sci. 2023 Nov 14;13(11):1589. doi: 10.3390/brainsci13111589. Brain Sci. 2023. PMID: 38002548 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials