

Recent advances in the development of protein-protein interactions modulators: mechanisms and clinical trials
- PMID: 32968059
- PMCID: PMC7511340
- DOI: 10.1038/s41392-020-00315-3
Recent advances in the development of protein-protein interactions modulators: mechanisms and clinical trials
Abstract
Protein-protein interactions (PPIs) have pivotal roles in life processes. The studies showed that aberrant PPIs are associated with various diseases, including cancer, infectious diseases, and neurodegenerative diseases. Therefore, targeting PPIs is a direction in treating diseases and an essential strategy for the development of new drugs. In the past few decades, the modulation of PPIs has been recognized as one of the most challenging drug discovery tasks. In recent years, some PPIs modulators have entered clinical studies, some of which been approved for marketing, indicating that the modulators targeting PPIs have broad prospects. Here, we summarize the recent advances in PPIs modulators, including small molecules, peptides, and antibodies, hoping to provide some guidance to the design of novel drugs targeting PPIs in the future.
Conflict of interest statement
The authors declare no competing interests.
Figures
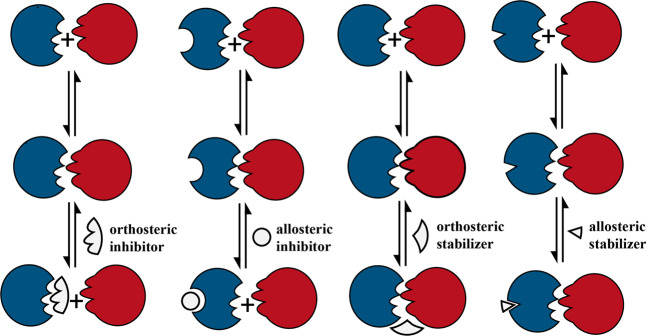
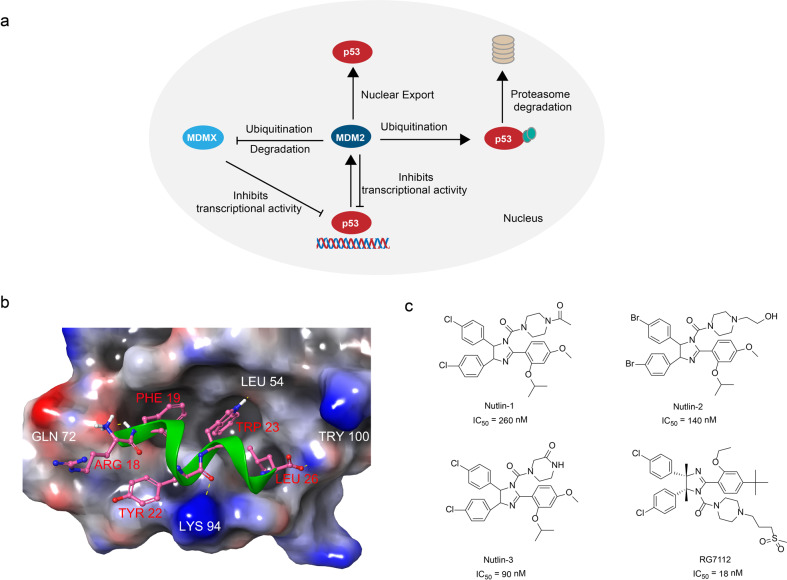
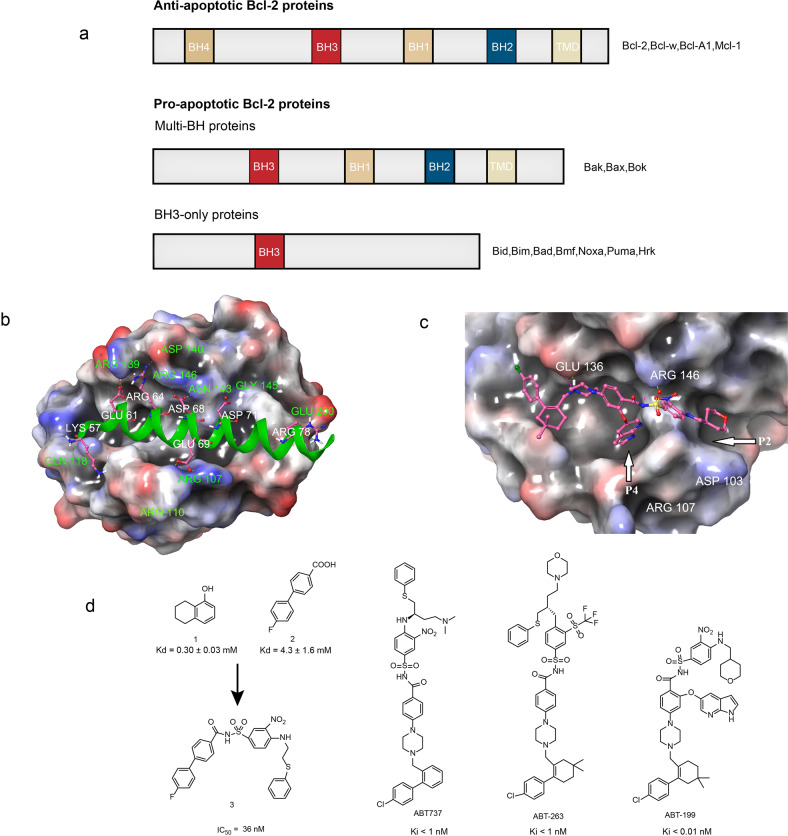
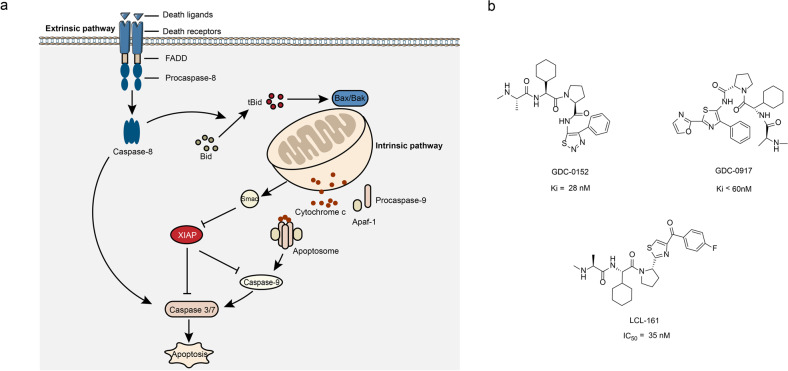
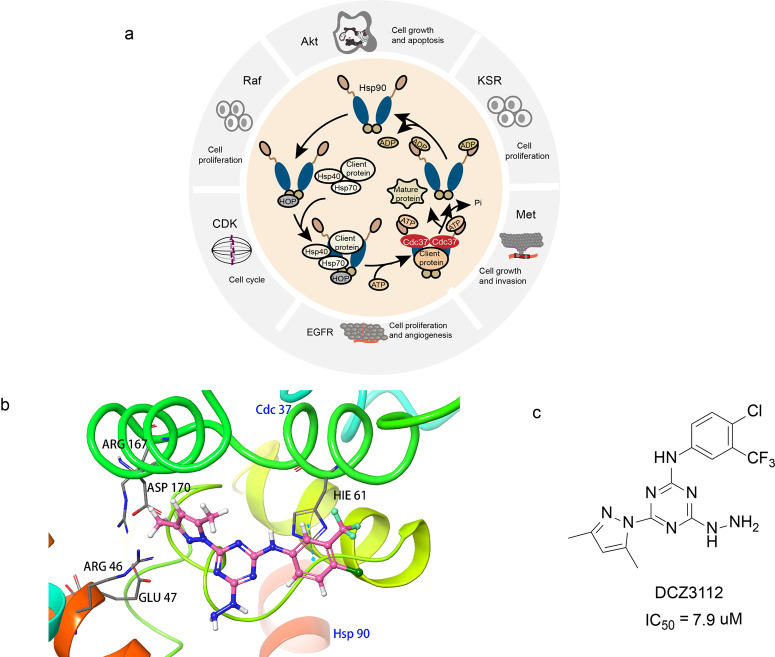
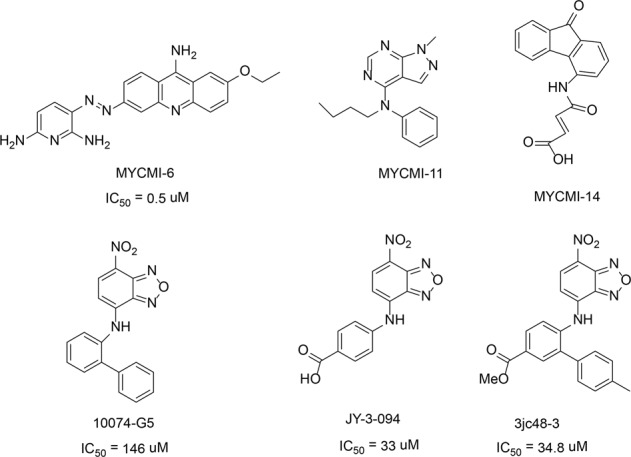
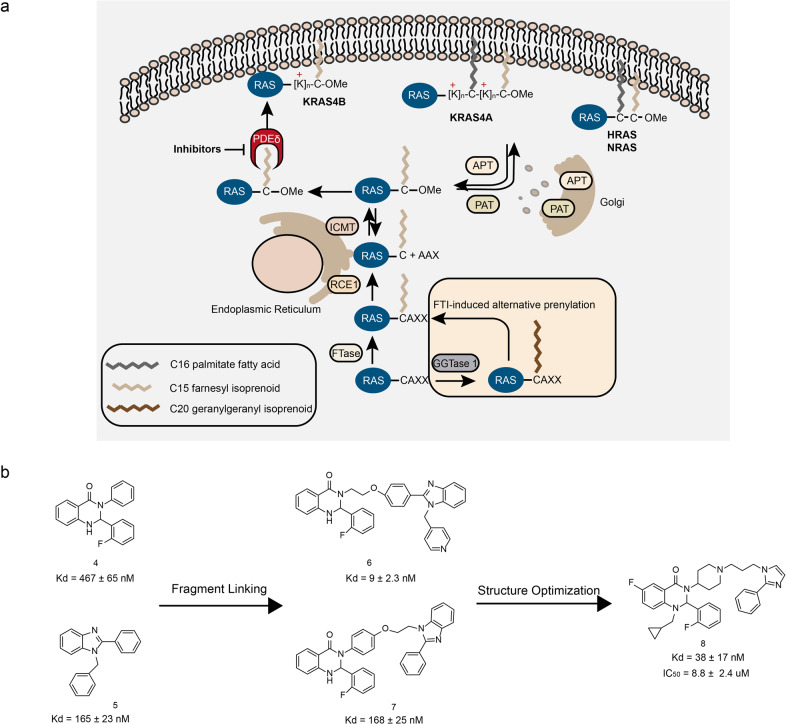
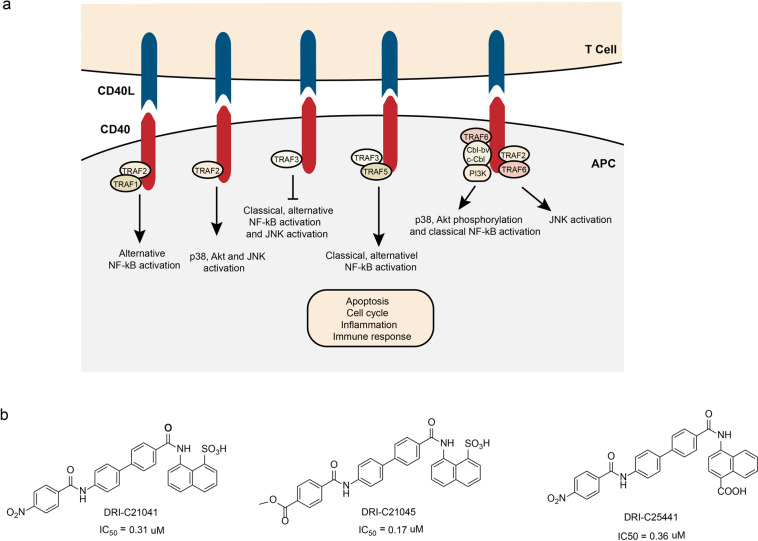
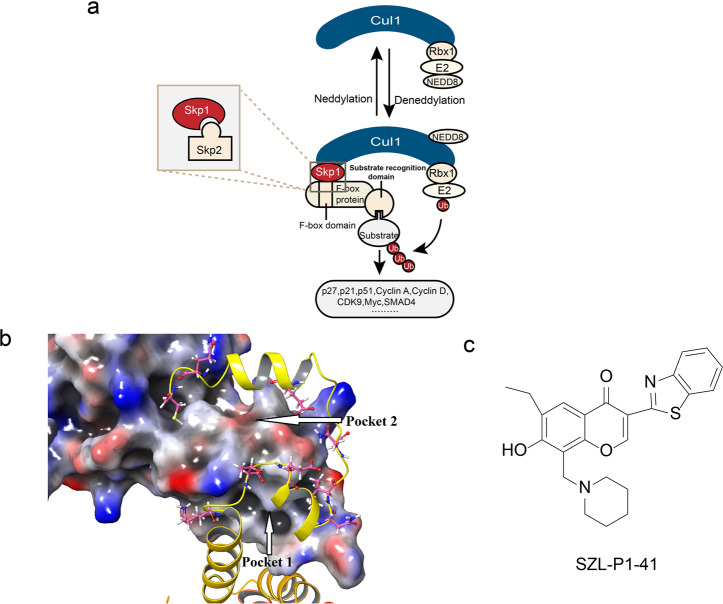
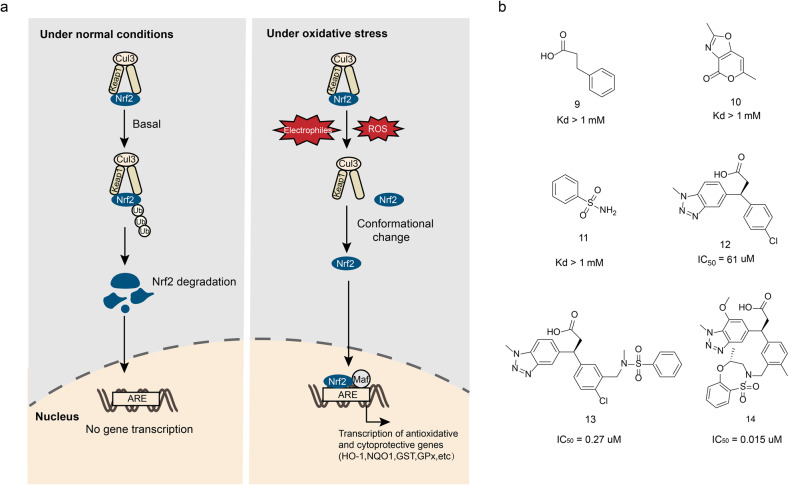
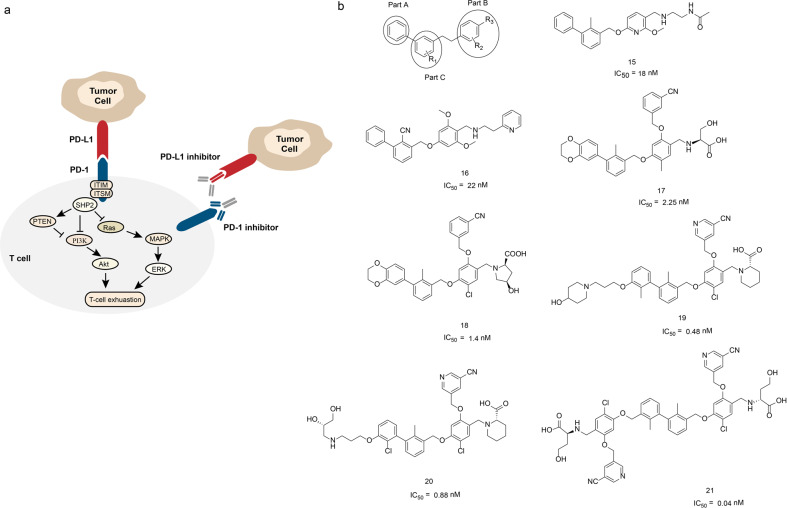
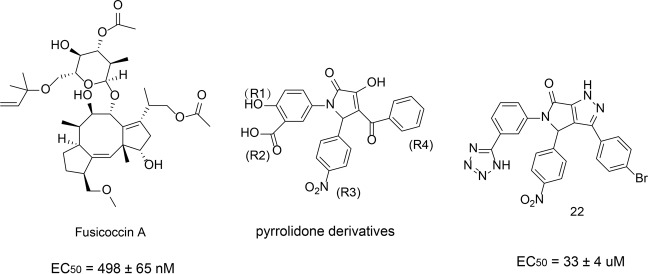
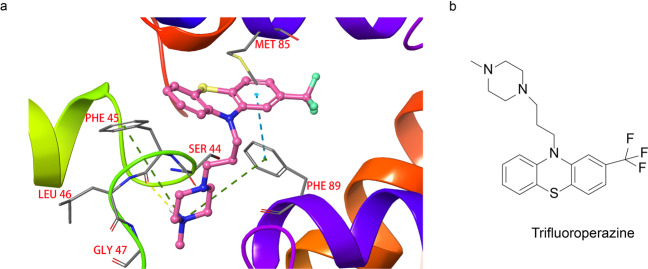
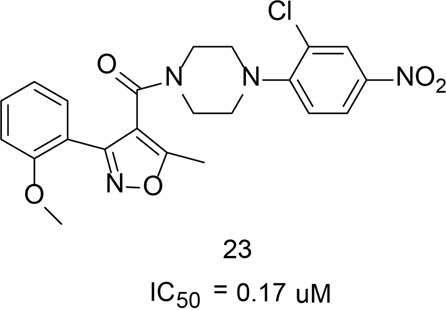
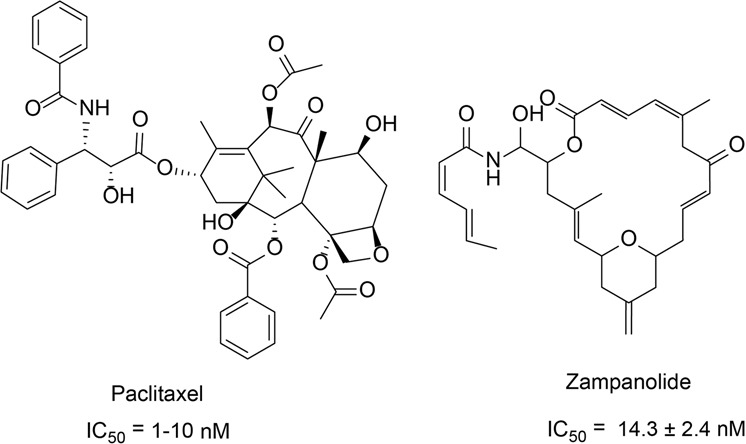
References
-
- Ferrari, S., Pellati, F. & Costi, M. P. Disruption of Protein-Protein Interfaces (Springer, Berlin, 2013).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
