

ATR is essential for preservation of cell mechanics and nuclear integrity during interstitial migration
- PMID: 32973141
- PMCID: PMC7518249
- DOI: 10.1038/s41467-020-18580-9
ATR is essential for preservation of cell mechanics and nuclear integrity during interstitial migration
Abstract
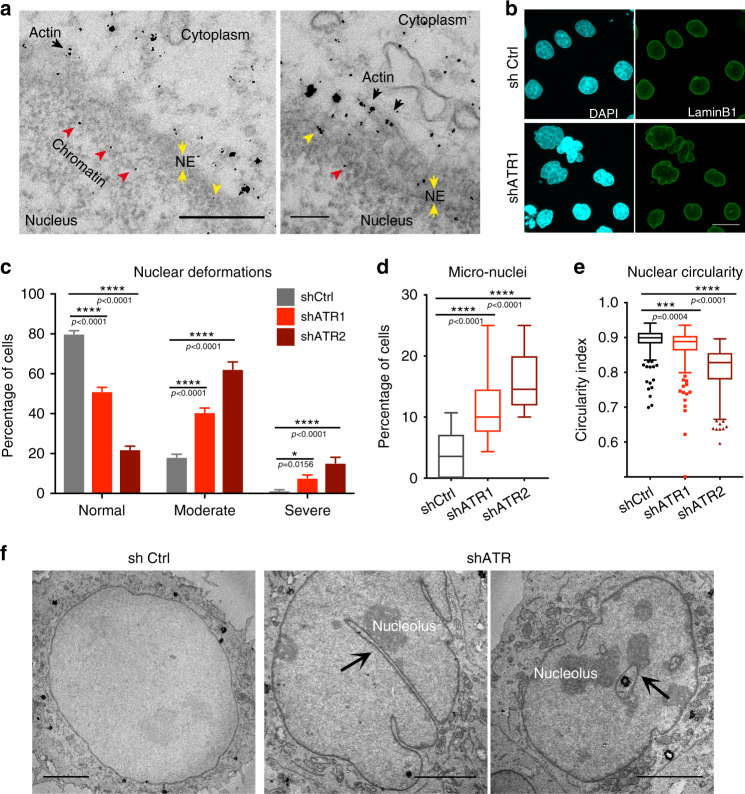
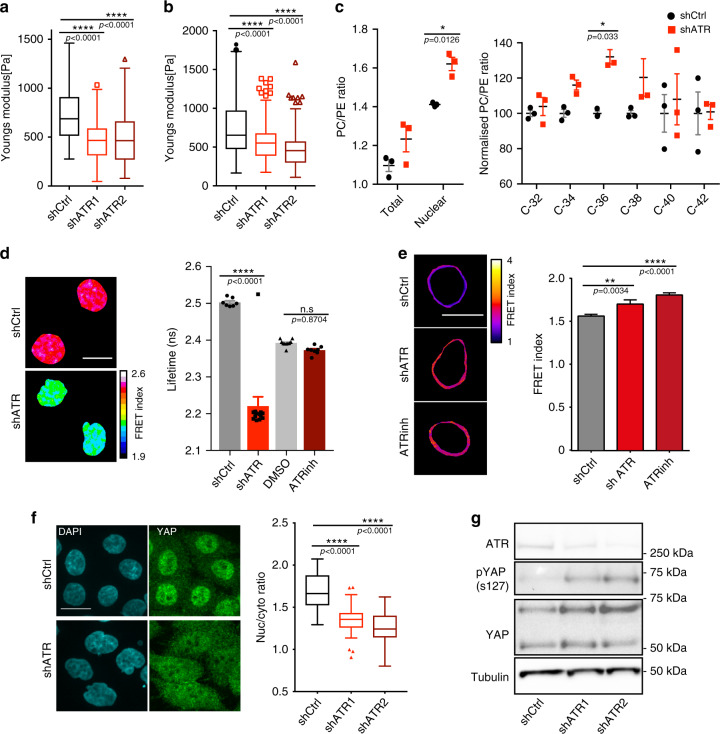
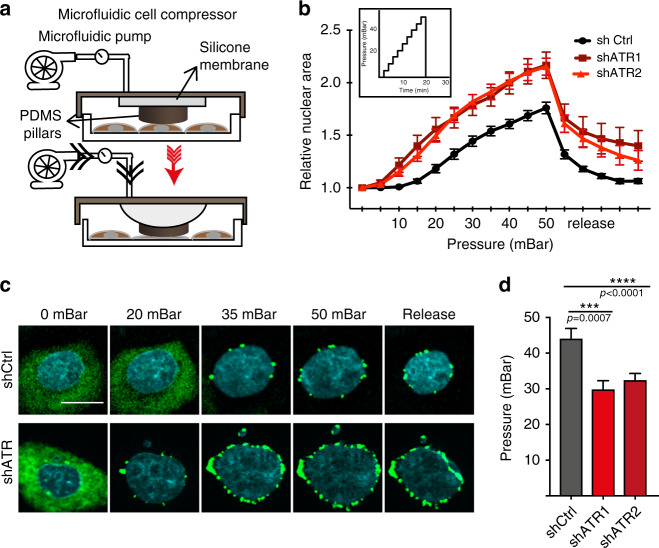
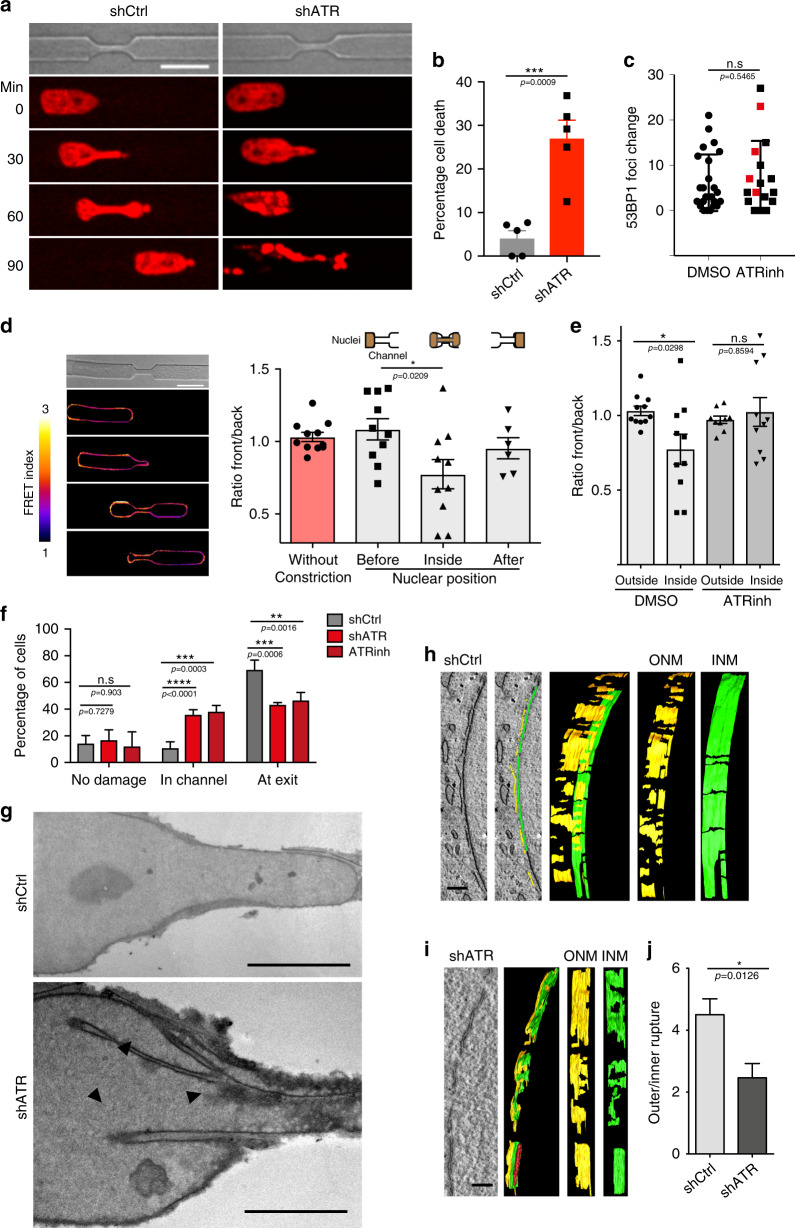
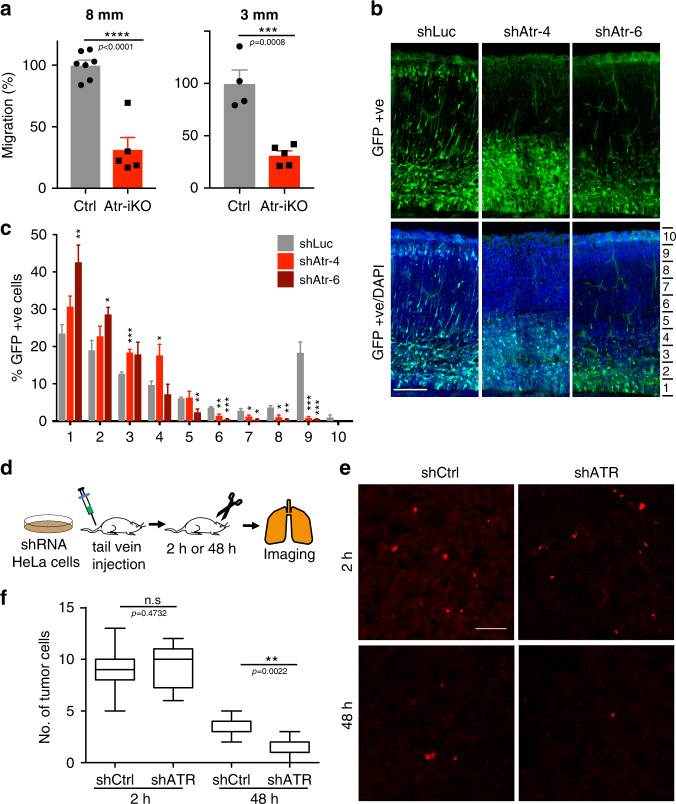
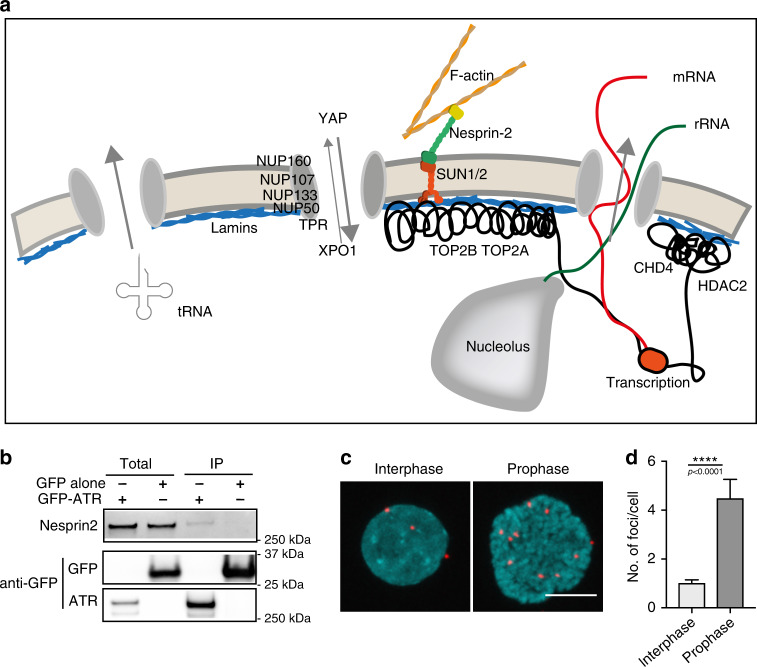
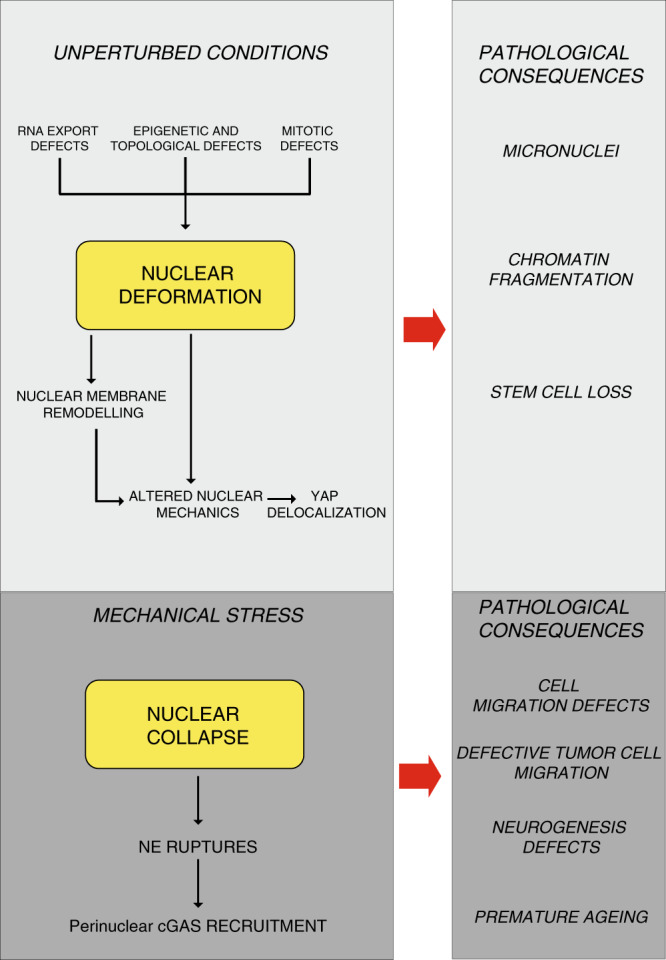
ATR responds to mechanical stress at the nuclear envelope and mediates envelope-associated repair of aberrant topological DNA states. By combining microscopy, electron microscopic analysis, biophysical and in vivo models, we report that ATR-defective cells exhibit altered nuclear plasticity and YAP delocalization. When subjected to mechanical stress or undergoing interstitial migration, ATR-defective nuclei collapse accumulating nuclear envelope ruptures and perinuclear cGAS, which indicate loss of nuclear envelope integrity, and aberrant perinuclear chromatin status. ATR-defective cells also are defective in neuronal migration during development and in metastatic dissemination from circulating tumor cells. Our findings indicate that ATR ensures mechanical coupling of the cytoskeleton to the nuclear envelope and accompanying regulation of envelope-chromosome association. Thus the repertoire of ATR-regulated biological processes extends well beyond its canonical role in triggering biochemical implementation of the DNA damage response.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
ATR-hippo drives force signaling to nuclear F-actin and links mechanotransduction to neurological disorders.Sci Adv. 2025 Feb 14;11(7):eadr5683. doi: 10.1126/sciadv.adr5683. Epub 2025 Feb 14. Sci Adv. 2025. PMID: 39951537 Free PMC article.
-
ATR mediates a checkpoint at the nuclear envelope in response to mechanical stress.Cell. 2014 Jul 31;158(3):633-46. doi: 10.1016/j.cell.2014.05.046. Cell. 2014. PMID: 25083873 Free PMC article.
-
Cell stretching activates an ATM mechano-transduction pathway that remodels cytoskeleton and chromatin.Cell Rep. 2023 Dec 26;42(12):113555. doi: 10.1016/j.celrep.2023.113555. Epub 2023 Dec 11. Cell Rep. 2023. PMID: 38088930
-
ATR-mediated regulation of nuclear and cellular plasticity.DNA Repair (Amst). 2016 Aug;44:143-150. doi: 10.1016/j.dnarep.2016.05.020. Epub 2016 May 16. DNA Repair (Amst). 2016. PMID: 27283761 Free PMC article. Review.
-
Regulation of Nuclear Mechanics and the Impact on DNA Damage.Int J Mol Sci. 2021 Mar 20;22(6):3178. doi: 10.3390/ijms22063178. Int J Mol Sci. 2021. PMID: 33804722 Free PMC article. Review.
Cited by
-
A TRilogy of ATR's Non-Canonical Roles Throughout the Cell Cycle and Its Relation to Cancer.Cancers (Basel). 2024 Oct 19;16(20):3536. doi: 10.3390/cancers16203536. Cancers (Basel). 2024. PMID: 39456630 Free PMC article. Review.
-
Small molecule metabolites: discovery of biomarkers and therapeutic targets.Signal Transduct Target Ther. 2023 Mar 20;8(1):132. doi: 10.1038/s41392-023-01399-3. Signal Transduct Target Ther. 2023. PMID: 36941259 Free PMC article. Review.
-
ATR inhibition enables complete tumour regression in ALK-driven NB mouse models.Nat Commun. 2021 Nov 24;12(1):6813. doi: 10.1038/s41467-021-27057-2. Nat Commun. 2021. PMID: 34819497 Free PMC article.
-
Nuclear morphology predicts cell survival to cisplatin chemotherapy.Neoplasia. 2023 Aug;42:100906. doi: 10.1016/j.neo.2023.100906. Epub 2023 May 10. Neoplasia. 2023. PMID: 37172462 Free PMC article.
-
ATR-hippo drives force signaling to nuclear F-actin and links mechanotransduction to neurological disorders.Sci Adv. 2025 Feb 14;11(7):eadr5683. doi: 10.1126/sciadv.adr5683. Epub 2025 Feb 14. Sci Adv. 2025. PMID: 39951537 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
