

A possible universal role for mRNA secondary structure in bacterial translation revealed using a synthetic operon
- PMID: 32973167
- PMCID: PMC7518266
- DOI: 10.1038/s41467-020-18577-4
A possible universal role for mRNA secondary structure in bacterial translation revealed using a synthetic operon
Abstract
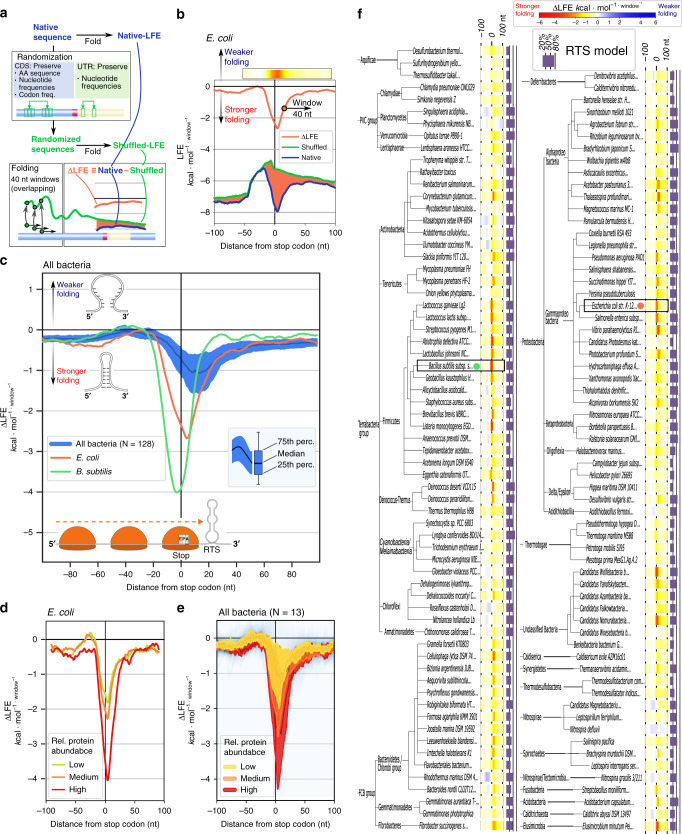
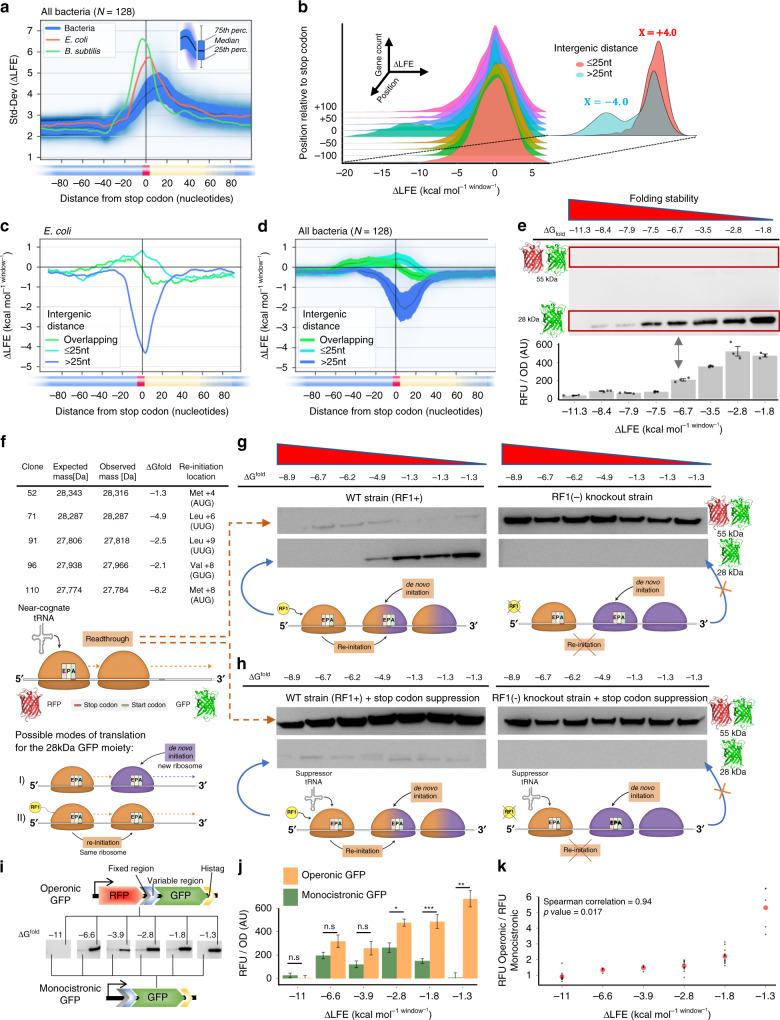
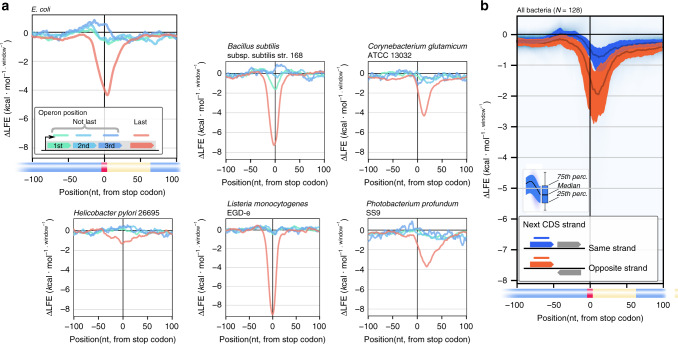
In bacteria, translation re-initiation is crucial for synthesizing proteins encoded by genes that are organized into operons. The mechanisms regulating translation re-initiation remain, however, poorly understood. We now describe the ribosome termination structure (RTS), a conserved and stable mRNA secondary structure localized immediately downstream of stop codons, and provide experimental evidence for its role in governing re-initiation efficiency in a synthetic Escherichia coli operon. We further report that RTSs are abundant, being associated with 18%-65% of genes in 128 analyzed bacterial genomes representing all phyla, and are selectively depleted when translation re-initiation is advantageous yet selectively enriched so as to insulate translation when re-initiation is deleterious. Our results support a potentially universal role for the RTS in controlling translation termination-insulation and re-initiation across bacteria.
Conflict of interest statement
T.T., L.A., Y.C. and M.P. have submitted a US provisional patent application regarding the use of synthetic RTSs to improve protein expression yields and control translation re-initiation. U.S. Provisional Patent: BGU-RMT-P-095-USP, Status: Pending. The remaining authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
