

A tunable L-arabinose-inducible expression plasmid for the acetic acid bacterium Gluconobacter oxydans
- PMID: 32974745
- PMCID: PMC7567684
- DOI: 10.1007/s00253-020-10905-4
A tunable L-arabinose-inducible expression plasmid for the acetic acid bacterium Gluconobacter oxydans
Abstract
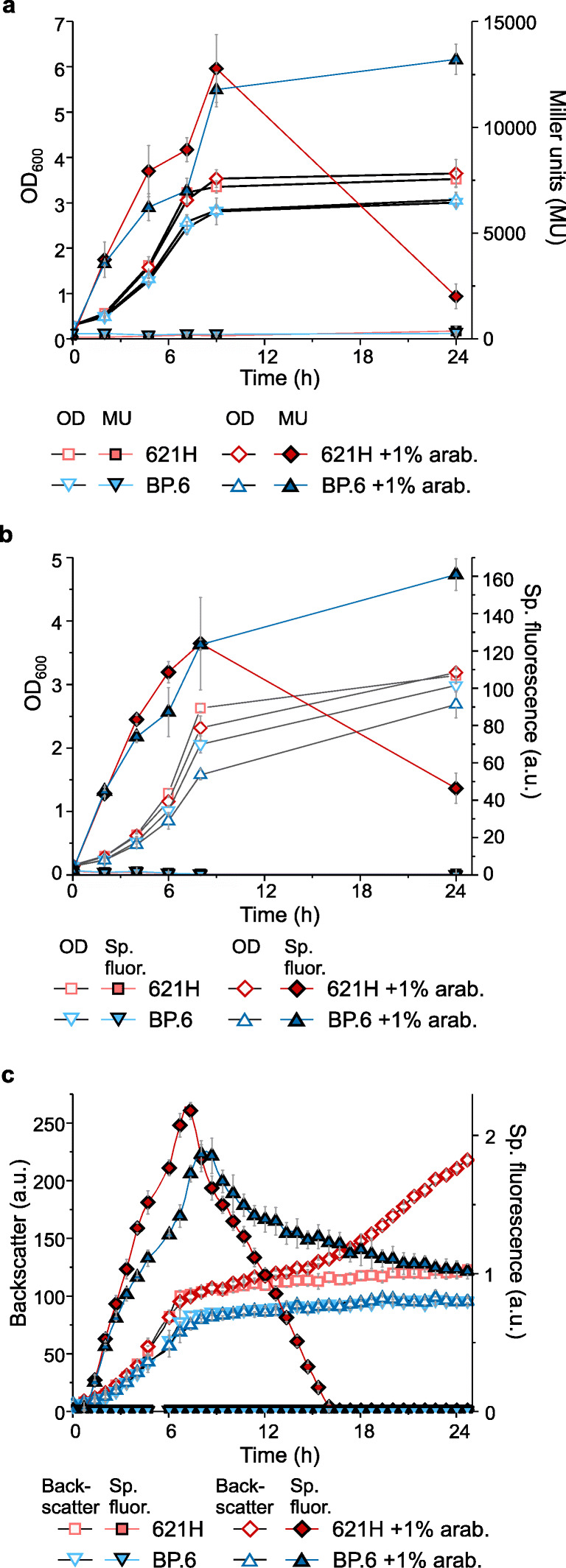
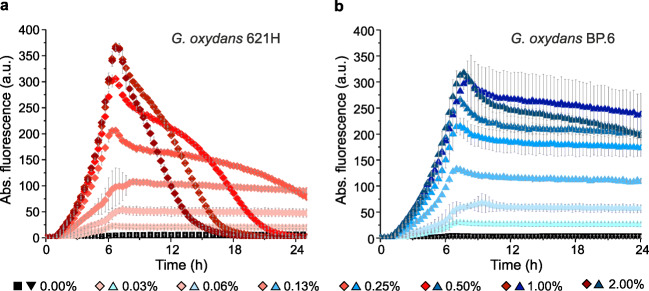
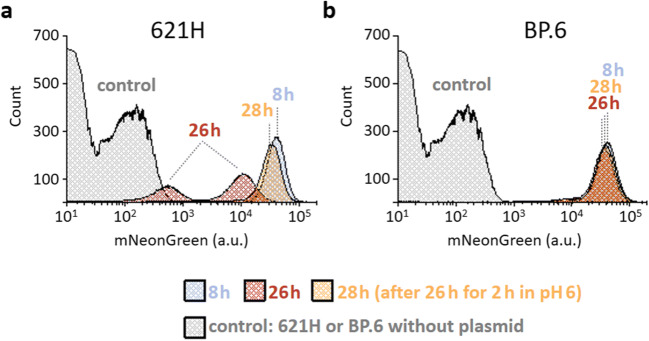
The acetic acid bacterium (AAB) Gluconobacter oxydans incompletely oxidizes a wide variety of carbohydrates and is therefore used industrially for oxidative biotransformations. For G. oxydans, no system was available that allows regulatable plasmid-based expression. We found that the L-arabinose-inducible PBAD promoter and the transcriptional regulator AraC from Escherichia coli MC4100 performed very well in G. oxydans. The respective pBBR1-based plasmids showed very low basal expression of the reporters β-glucuronidase and mNeonGreen, up to 480-fold induction with 1% L-arabinose, and tunability from 0.1 to 1% L-arabinose. In G. oxydans 621H, L-arabinose was oxidized by the membrane-bound glucose dehydrogenase, which is absent in the multi-deletion strain BP.6. Nevertheless, AraC-PBAD performed similar in both strains in the exponential phase, indicating that a gene knockout is not required for application of AraC-PBAD in wild-type G. oxydans strains. However, the oxidation product arabinonic acid strongly contributed to the acidification of the growth medium in 621H cultures during the stationary phase, which resulted in drastically decreased reporter activities in 621H (pH 3.3) but not in BP.6 cultures (pH 4.4). These activities could be strongly increased quickly solely by incubating stationary cells in D-mannitol-free medium adjusted to pH 6, indicating that the reporters were hardly degraded yet rather became inactive. In a pH-controlled bioreactor, these reporter activities remained high in the stationary phase (pH 6). Finally, we created a multiple cloning vector with araC-PBAD based on pBBR1MCS-5. Together, we demonstrated superior functionality and good tunability of an AraC-PBAD system in G. oxydans that could possibly also be used in other AAB. KEY POINTS: • We found the AraC-PBAD system from E. coli MC4100 was well tunable in G. oxydans. • In the absence of AraC or l-arabinose, expression from PBAD was extremely low. • This araC-PBAD system could also be fully functional in other acetic acid bacteria.
Keywords: AraC; Induction; Membrane-bound dehydrogenase; PBAD promoter; mNeonGreen; β-D-Glucuronidase UidA.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
