

The Self-Organizing Genome: Principles of Genome Architecture and Function
- PMID: 32976797
- PMCID: PMC7541718
- DOI: 10.1016/j.cell.2020.09.014
The Self-Organizing Genome: Principles of Genome Architecture and Function
Abstract
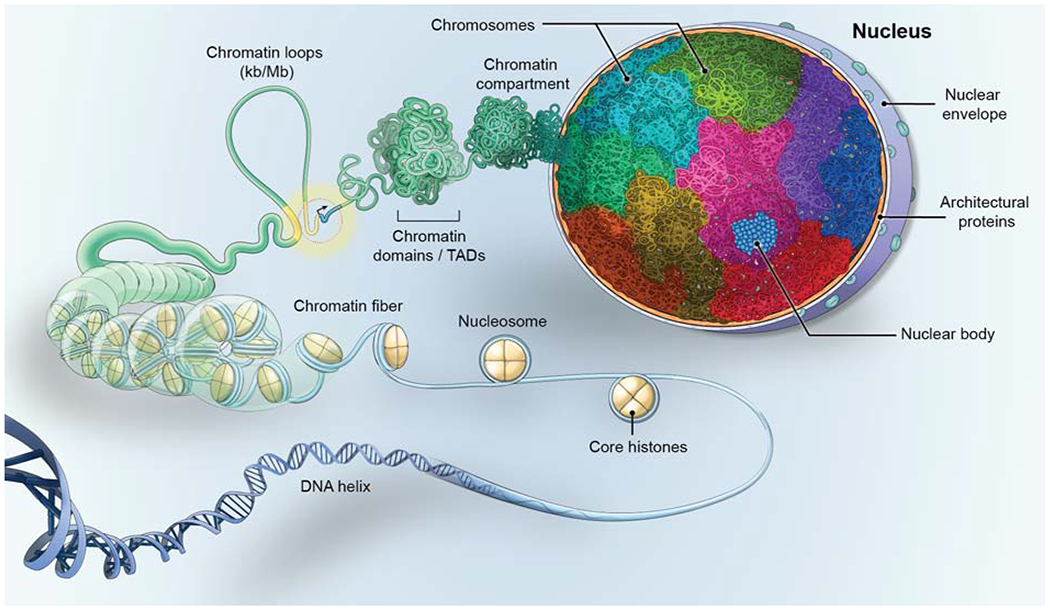
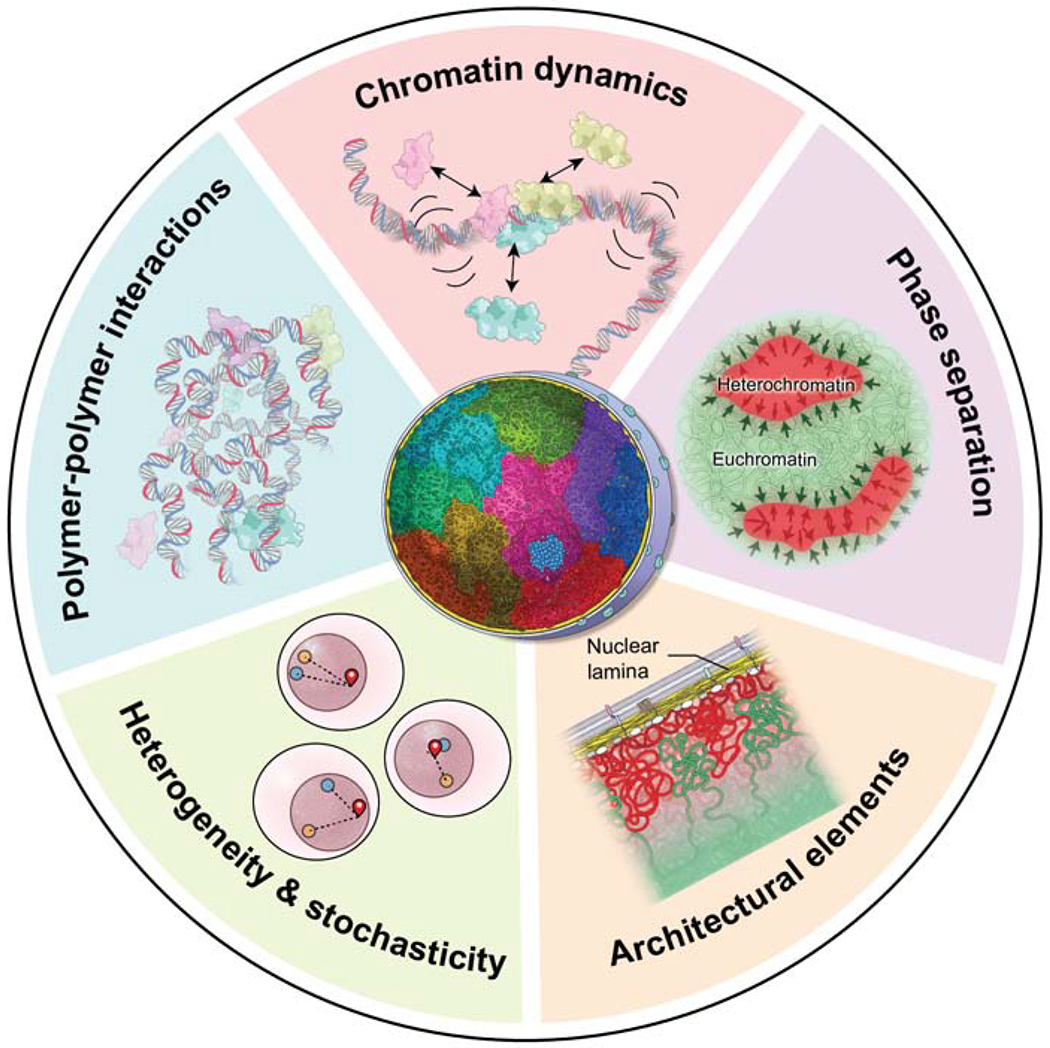
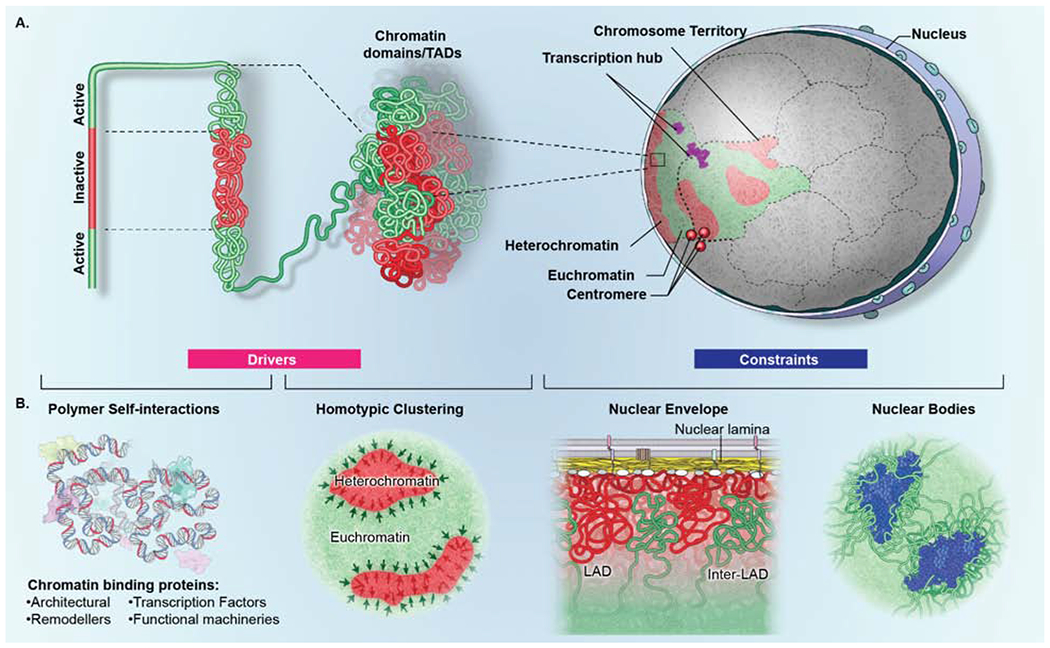
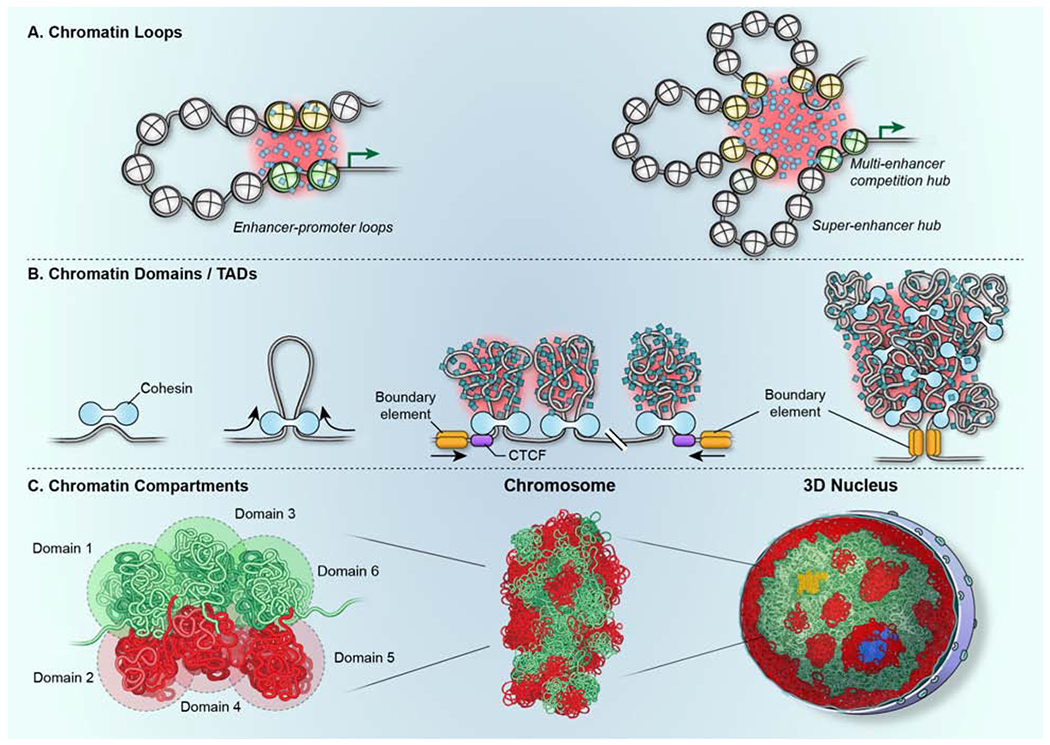
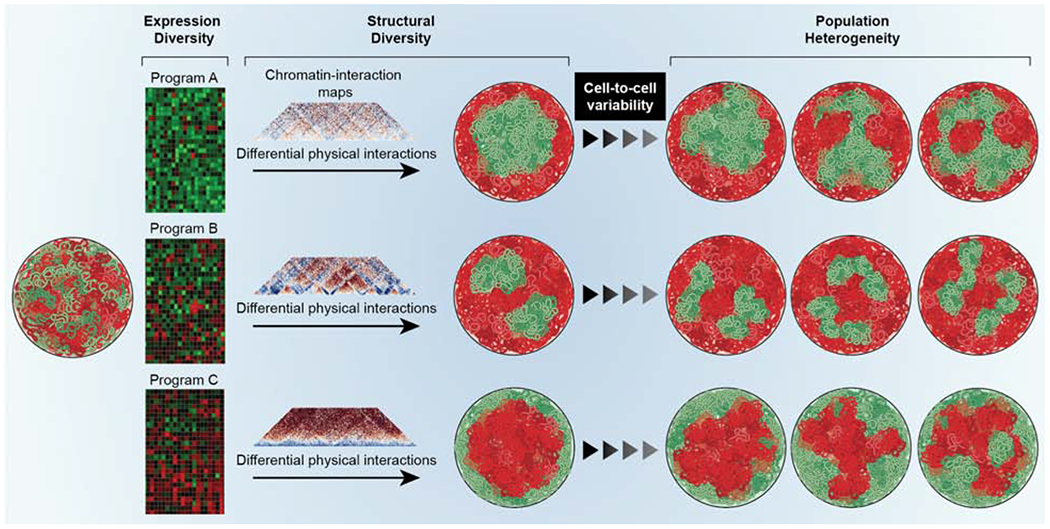
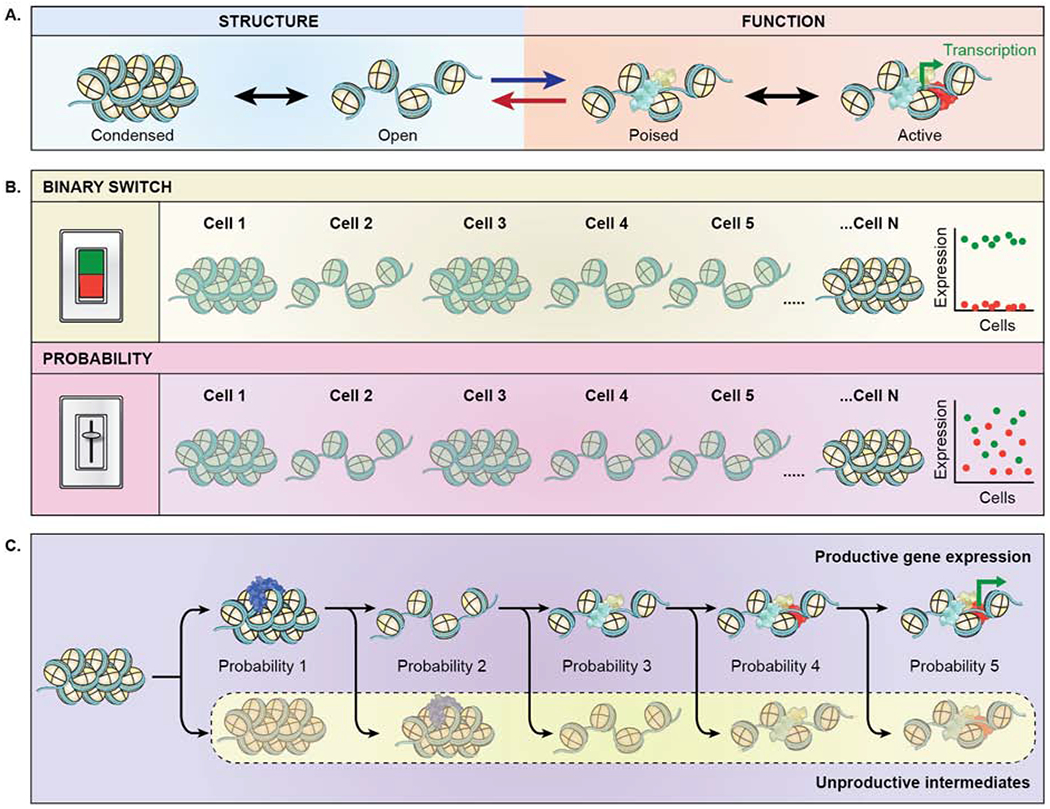
Genomes have complex three-dimensional architectures. The recent convergence of genetic, biochemical, biophysical, and cell biological methods has uncovered several fundamental principles of genome organization. They highlight that genome function is a major driver of genome architecture and that structural features of chromatin act as modulators, rather than binary determinants, of genome activity. The interplay of these principles in the context of self-organization can account for the emergence of structural chromatin features, the diversity and single-cell heterogeneity of nuclear architecture in cell types and tissues, and explains evolutionarily conserved functional features of genomes, including plasticity and robustness.
Keywords: chromatin; dynamics; gene expression; genome organization; nuclear architecture; phase separation; self-organization.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests
The author declares no competing interests.
Figures
References
-
- Bar-Even A, Paulsson J, Maheshri N, Carmi M, O’Shea E, Pilpel Y, and Barkai N (2006). Noise in protein expression scales with natural protein abundance. Nat Genet 38, 636–643. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
