

A Putative Bet-Hedging Strategy Buffers Budding Yeast against Environmental Instability
- PMID: 32976801
- PMCID: PMC7726042
- DOI: 10.1016/j.cub.2020.08.092
A Putative Bet-Hedging Strategy Buffers Budding Yeast against Environmental Instability
Abstract
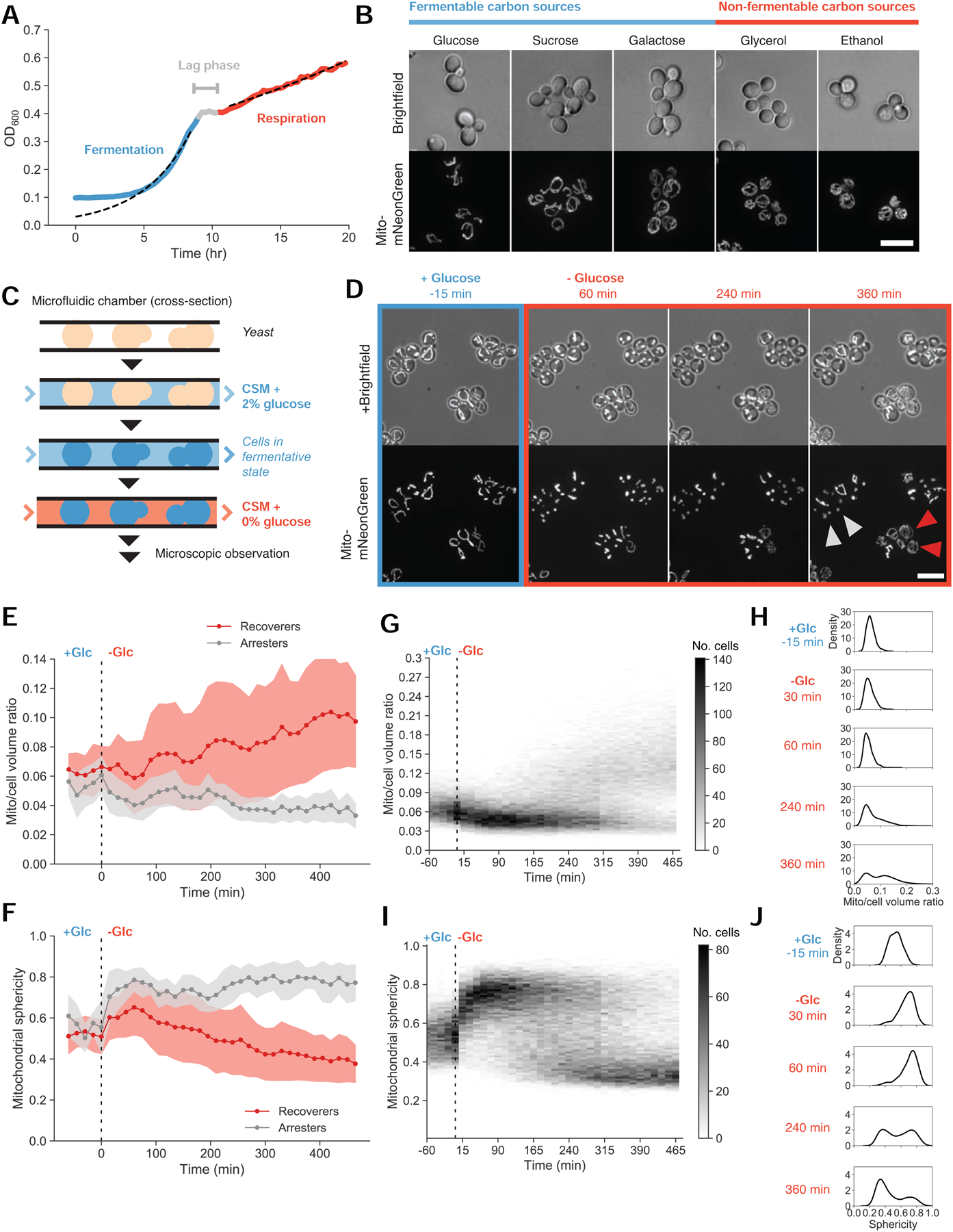
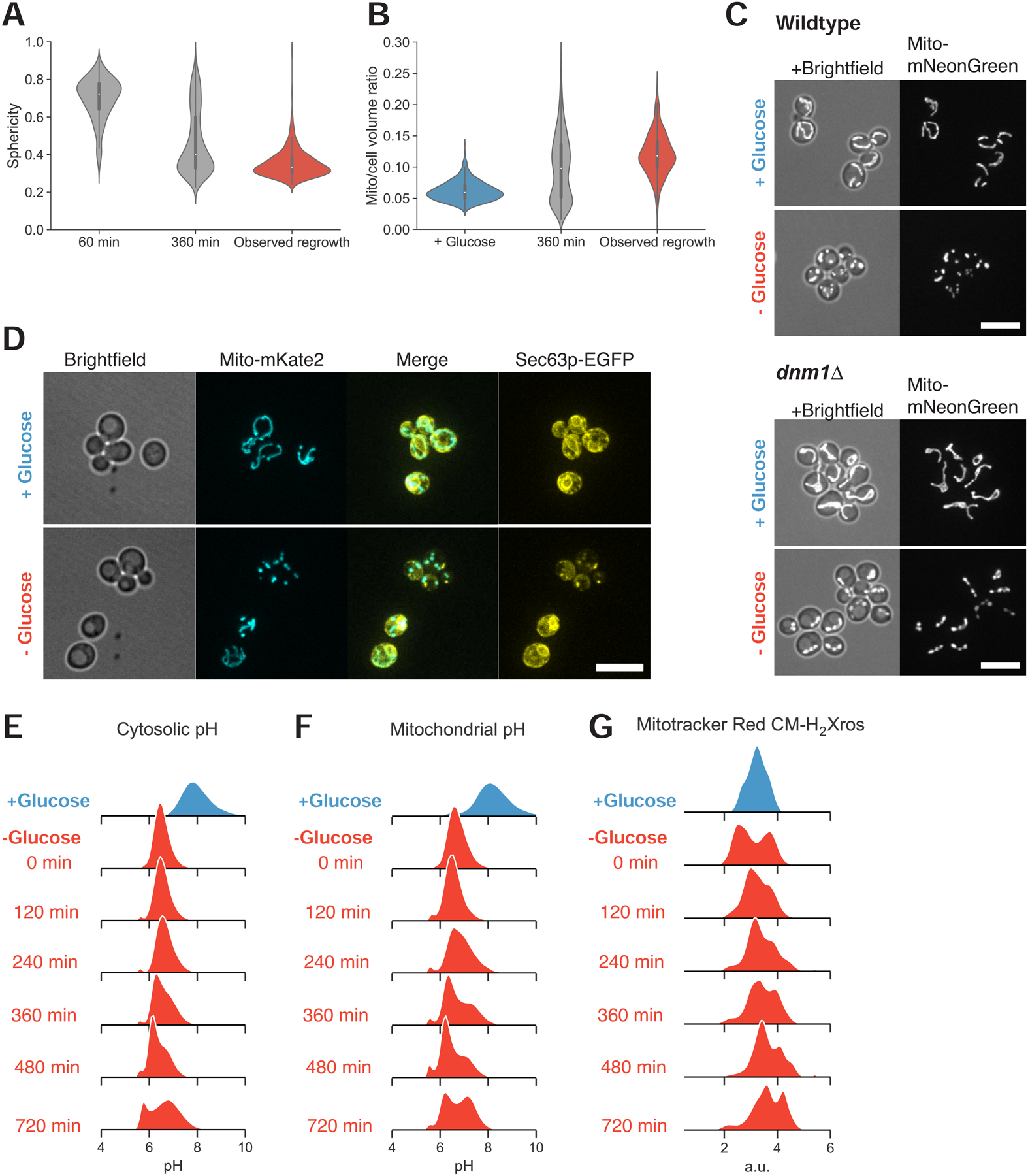
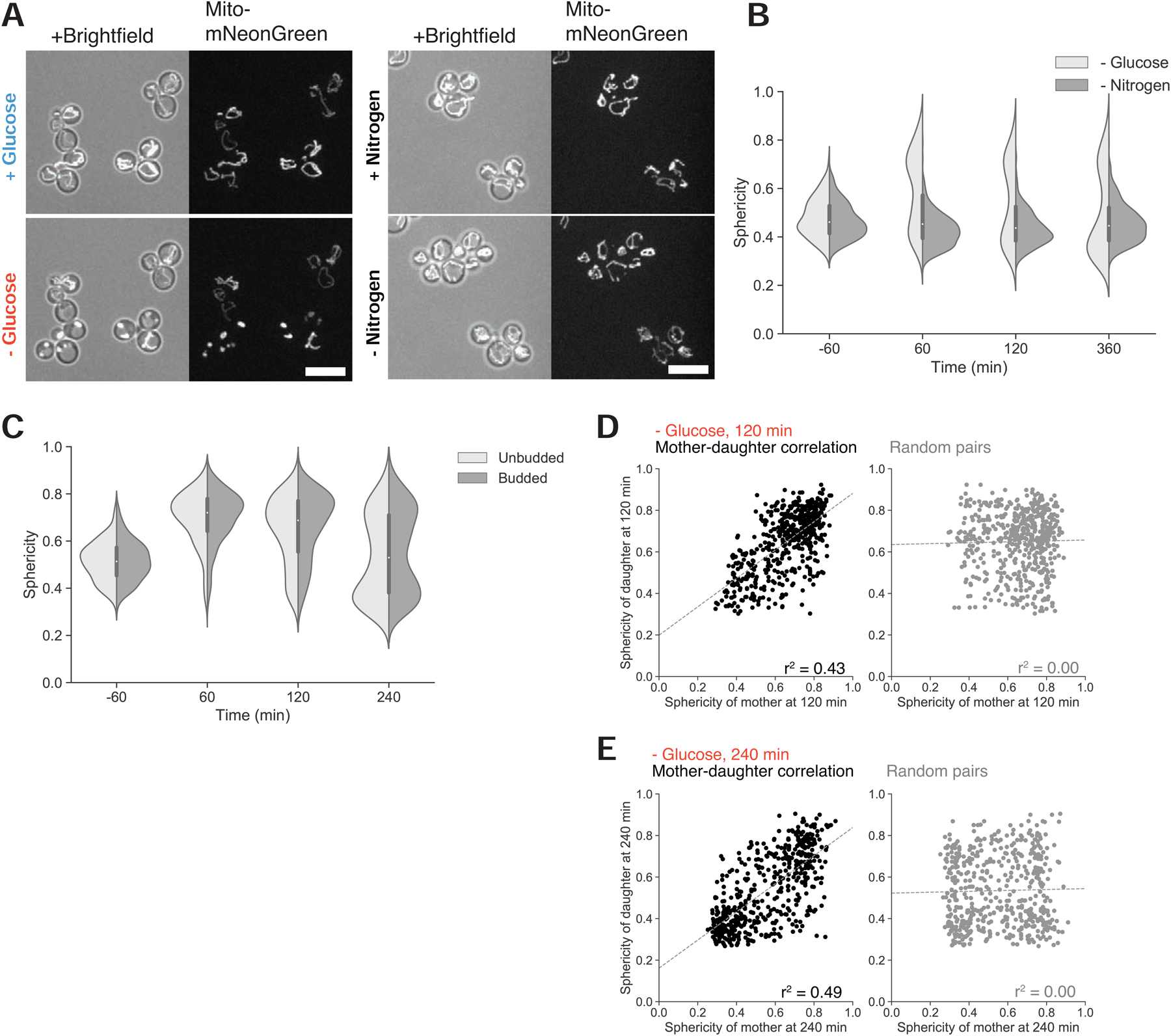
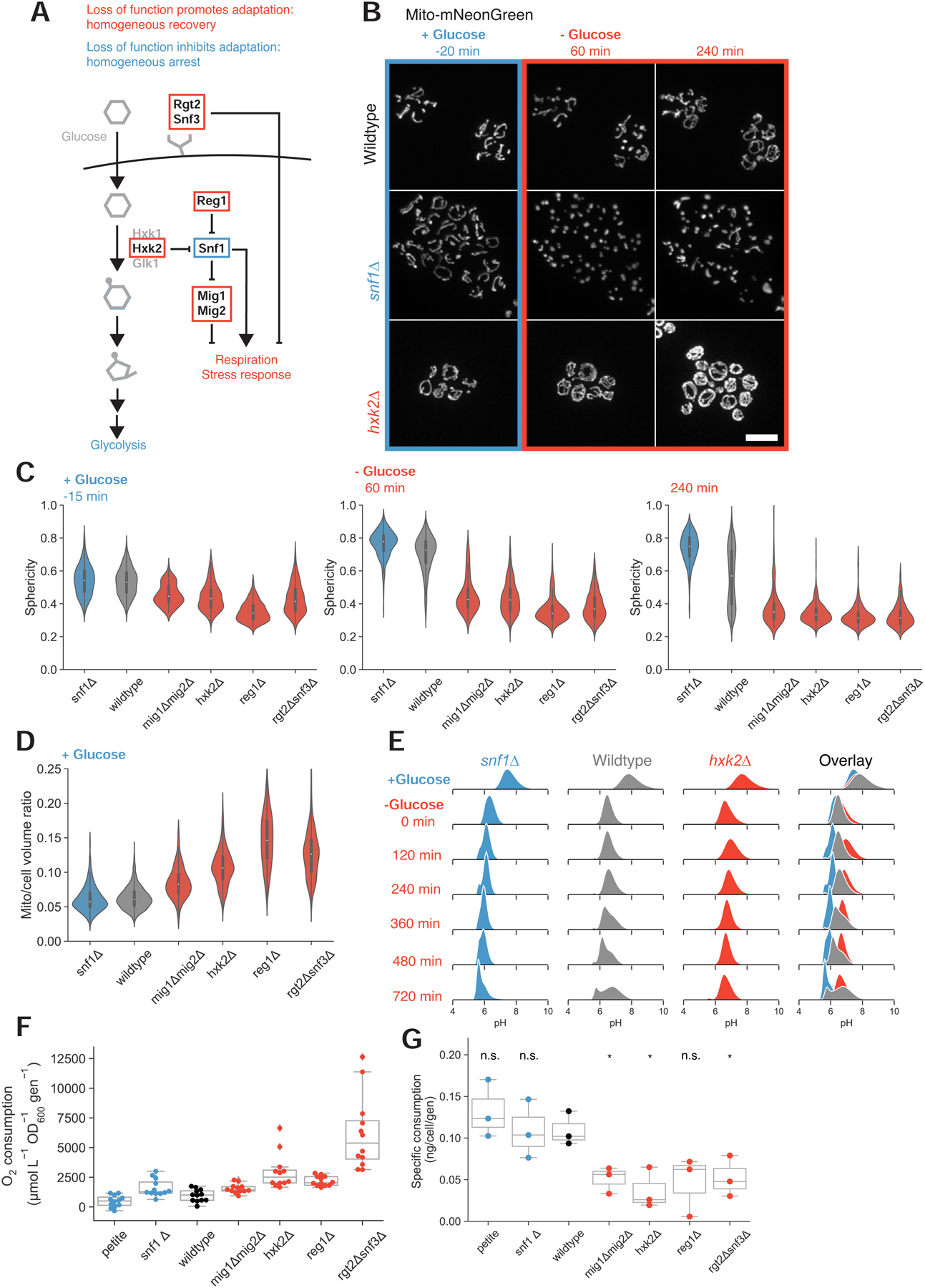
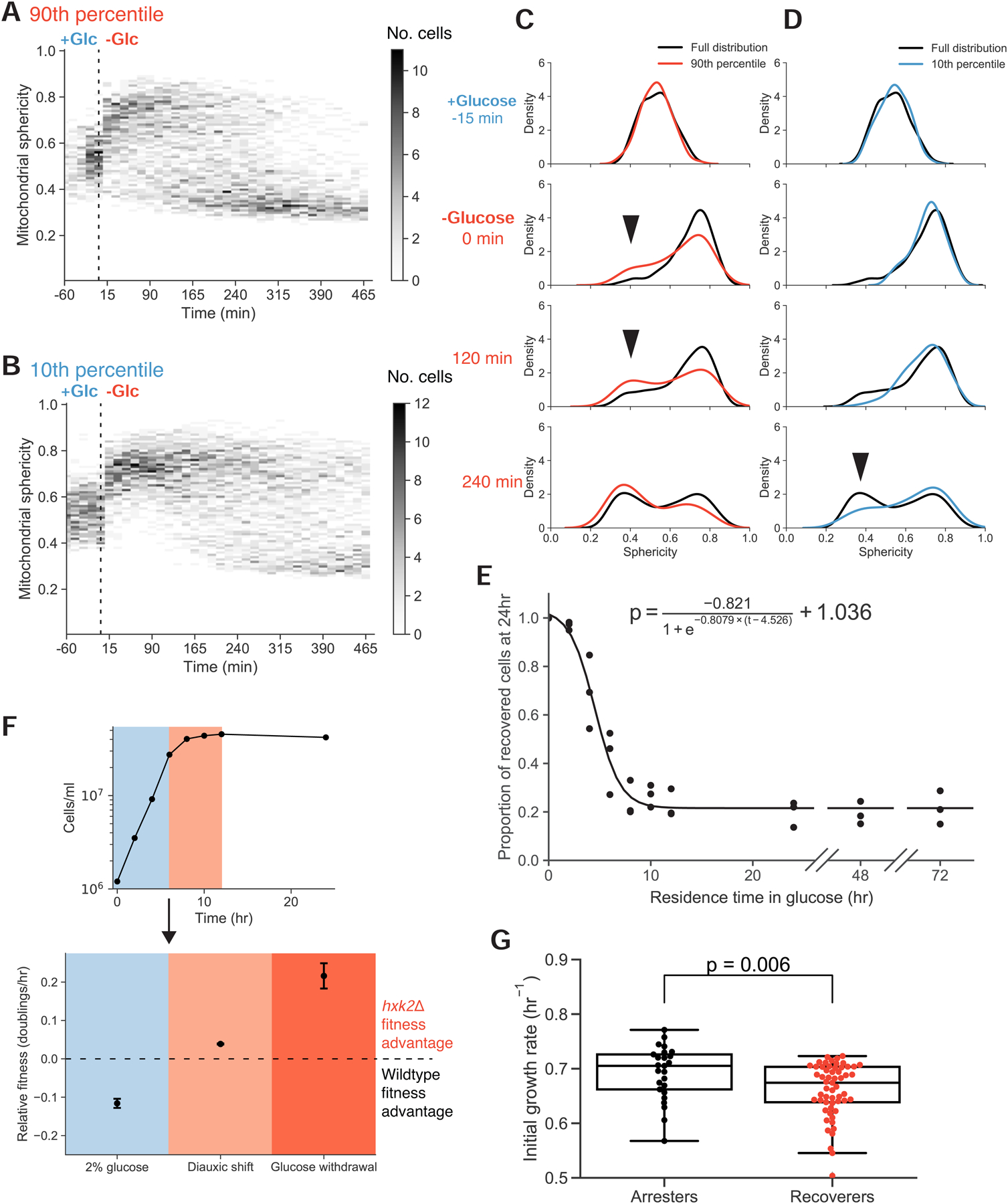
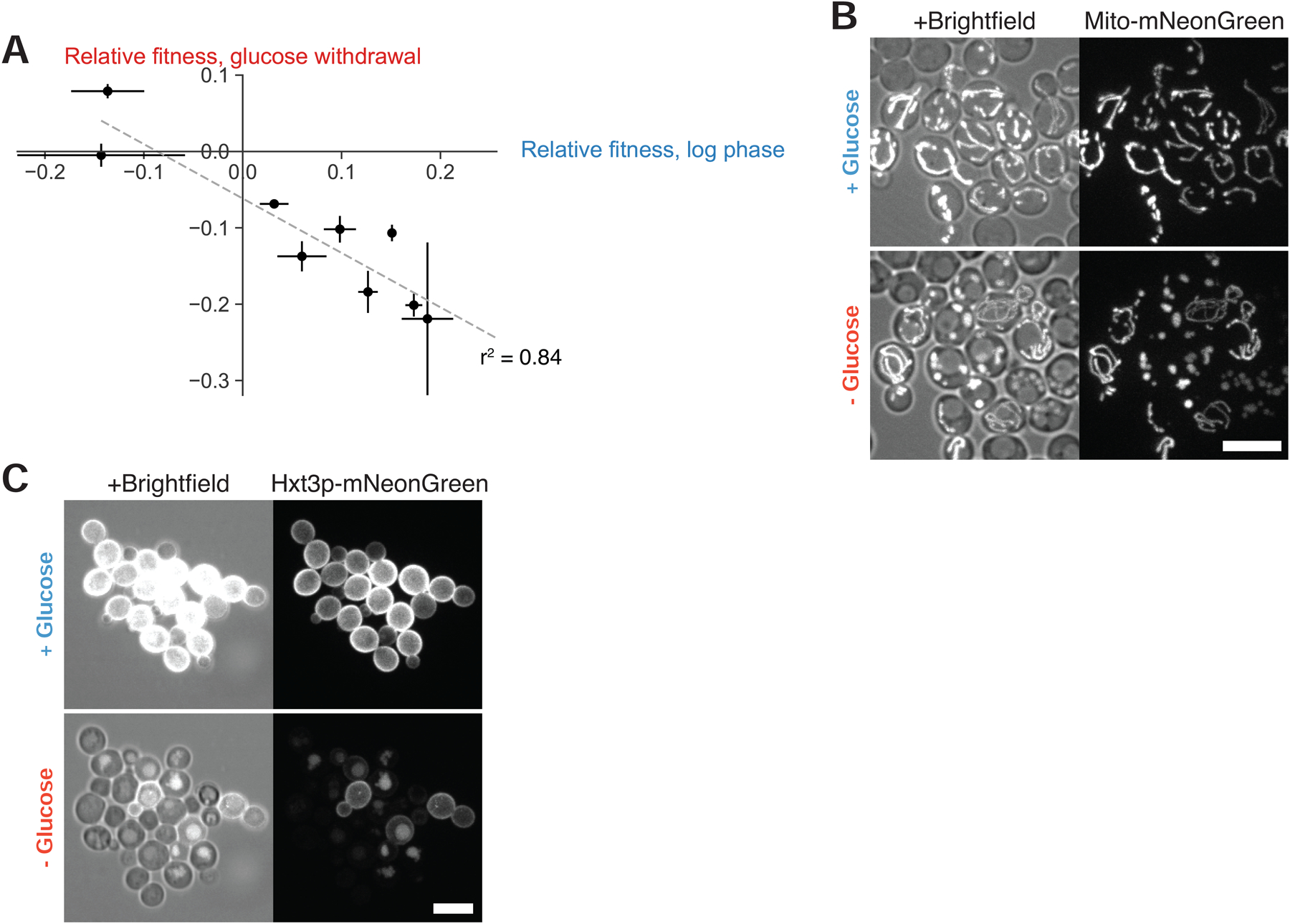
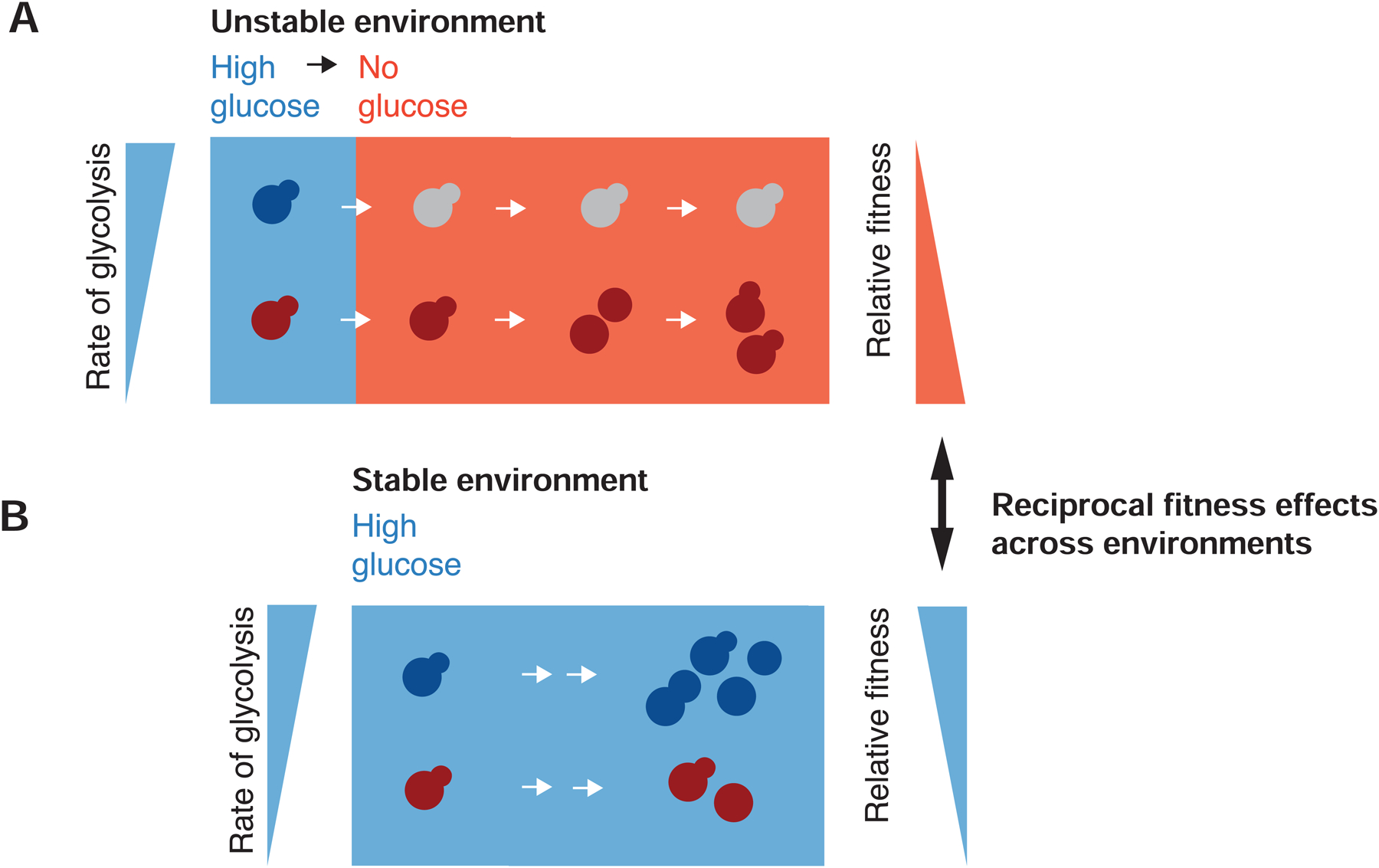
To grow and divide, cells must extract resources from dynamic and unpredictable environments. Many organisms use different metabolic strategies for distinct contexts. Budding yeast can produce ATP from carbon sources by mechanisms that prioritize either speed (fermentation) or yield (respiration). Withdrawing glucose from exponentially growing cells reveals variability in their ability to switch from fermentation to respiration. We observe two subpopulations of glucose-starved cells: recoverers, which rapidly adapt and resume growth, and arresters, which enter a shock state characterized by deformation of many cellular structures, including mitochondria. These states are heritable, and on high glucose, arresters grow and divide faster than recoverers. Recoverers have a fitness advantage during a carbon source shift but are less fit in a constant, high-glucose environment, and we observe natural variation in the frequency of the two states across wild yeast strains. These experiments suggest that bet hedging has evolved in budding yeast.
Keywords: bet hedging; bimodality; budding yeast; fermentation; metabolism; mitochondria; respiration.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Johnston M, and Carlson M (1992). Carbon regulation in Saccharomyces. Molecular and Cellular Biology of the Yeast Saccharomyces, Broach JR, Pringle JR, Jones EW (eds) Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
-
- Zaman S, Lippman SI, Zhao X, and Broach JR (2008). How Saccharomyces responds to nutrients. Annu. Rev. Genet 42, 27–81. - PubMed
-
- Yotsuyanagi Y (1962). [Study of yeast mitochondria. I. Variations in mitochondrial ultrastructure during the aerobic growth cycle]. J. Ultrastruct. Res 7, 121–140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
