

Efficiency of the four proteasome subtypes to degrade ubiquitinated or oxidized proteins
- PMID: 32978409
- PMCID: PMC7519072
- DOI: 10.1038/s41598-020-71550-5
Efficiency of the four proteasome subtypes to degrade ubiquitinated or oxidized proteins
Abstract
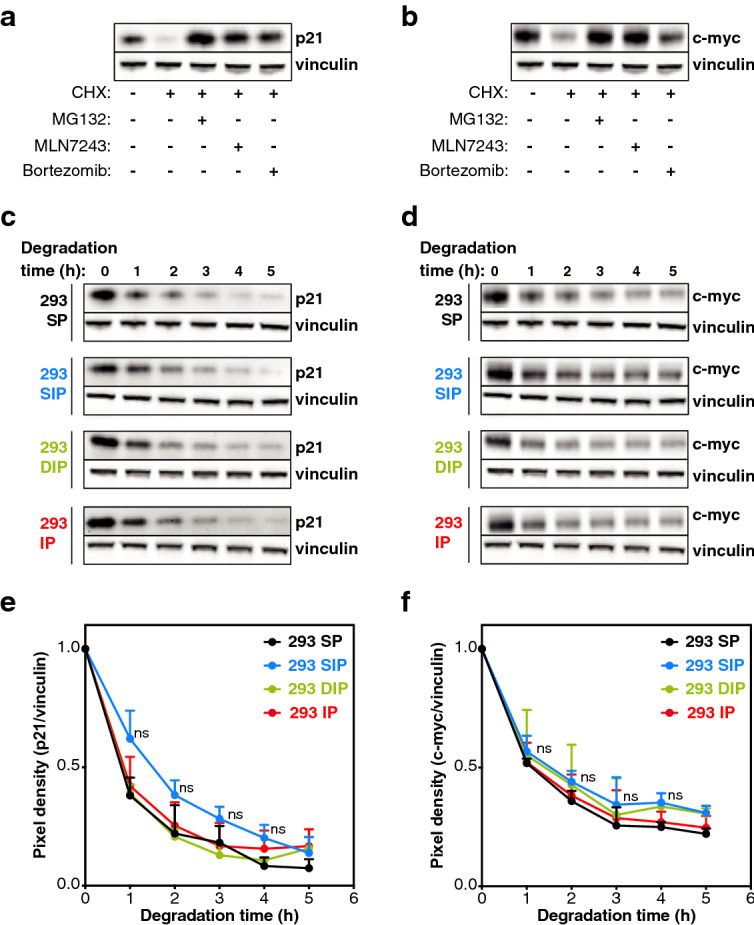
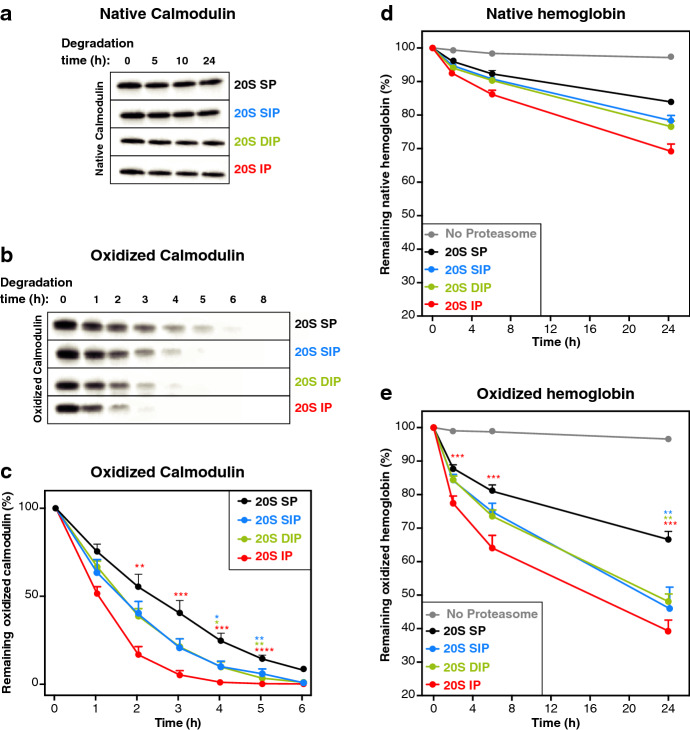
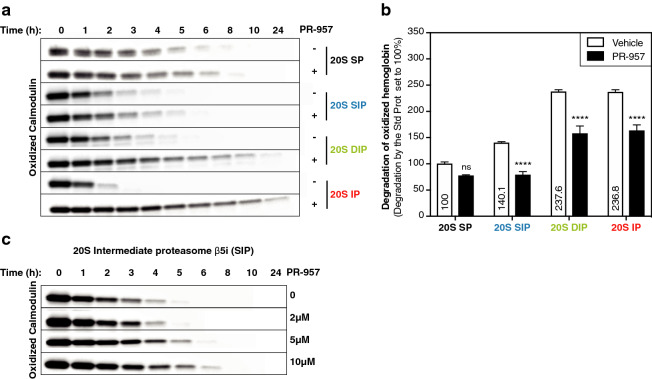
The proteasome is responsible for selective degradation of proteins. It exists in mammalian cells under four main subtypes, which differ by the combination of their catalytic subunits: the standard proteasome (β1-β2-β5), the immunoproteasome (β1i-β2i-β5i) and the two intermediate proteasomes (β1-β2-β5i and β1i-β2-β5i). The efficiency of the four proteasome subtypes to degrade ubiquitinated or oxidized proteins remains unclear. Using cells expressing exclusively one proteasome subtype, we observed that ubiquitinated p21 and c--myc were degraded at similar rates, indicating that the four 26S proteasomes degrade ubiquitinated proteins equally well. Under oxidative stress, we observed a partial dissociation of 26S into 20S proteasomes, which can degrade non-ubiquitinated oxidized proteins. Oxidized calmodulin and hemoglobin were best degraded in vitro by the three β5i-containing 20S proteasomes, while their native forms were not degraded. Circular dichroism analyses indicated that ubiquitin-independent recognition of oxidized proteins by 20S proteasomes was triggered by the disruption of their structure. Accordingly, β5i-containing 20S proteasomes degraded unoxidized naturally disordered protein tau, while 26S proteasomes did not. Our results suggest that the three β5i-containing 20S proteasomes, namely the immunoproteasome and the two intermediate proteasomes, might help cells to eliminate proteins containing disordered domains, including those induced by oxidative stress.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
