

The genomes of a monogenic fly: views of primitive sex chromosomes
- PMID: 32978490
- PMCID: PMC7519133
- DOI: 10.1038/s41598-020-72880-0
The genomes of a monogenic fly: views of primitive sex chromosomes
Abstract
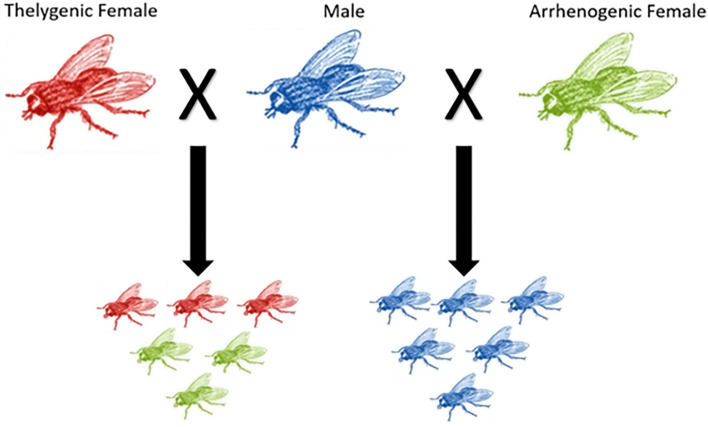
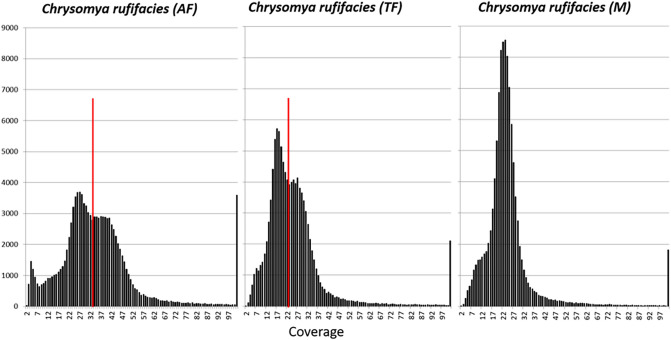
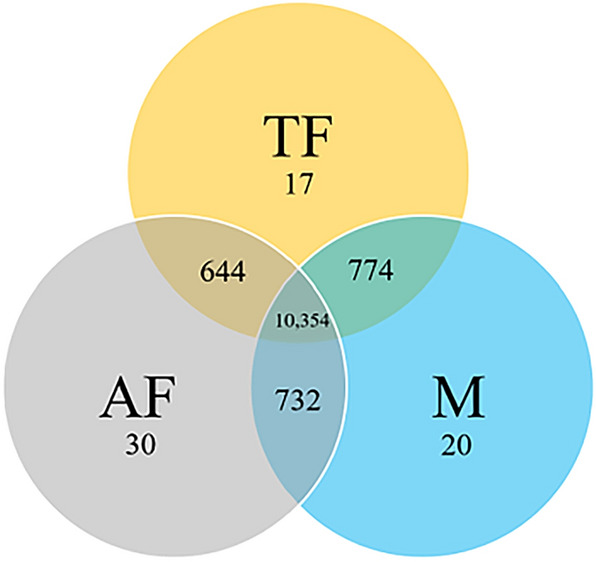
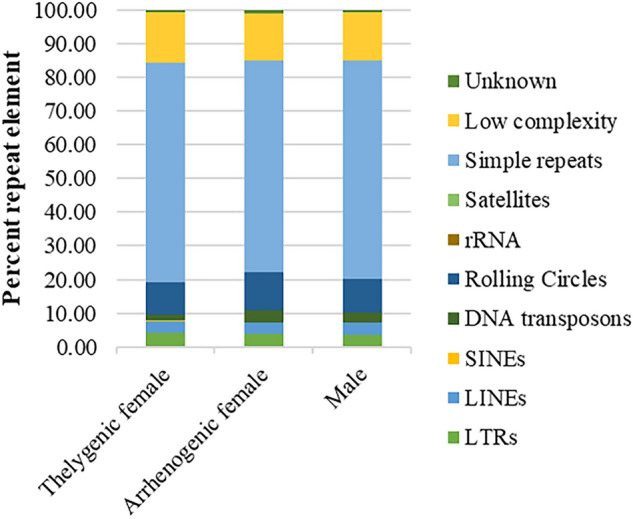
The production of male and female offspring is often determined by the presence of specific sex chromosomes which control sex-specific expression, and sex chromosomes evolve through reduced recombination and specialized gene content. Here we present the genomes of Chrysomya rufifacies, a monogenic blow fly (females produce female or male offspring, exclusively) by separately sequencing and assembling each type of female and the male. The genomes (> 25X coverage) do not appear to have any sex-linked Muller F elements (typical for many Diptera) and exhibit little differentiation between groups supporting the morphological assessments of C. rufifacies homomorphic chromosomes. Males in this species are associated with a unimodal coverage distribution while females exhibit bimodal coverage distributions, suggesting a potential difference in genomic architecture. The presence of the individual-sex draft genomes herein provides new clues regarding the origination and evolution of the diverse sex-determining mechanisms observed within Diptera. Additional genomic analysis of sex chromosomes and sex-determining genes of other blow flies will allow a refined evolutionary understanding of how flies with a typical X/Y heterogametic amphogeny (male and female offspring in similar ratios) sex determination systems evolved into one with a dominant factor that results in single sex progeny in a chromosomally monomorphic system.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
