

Functional analysis of Cti6 core domain responsible for recruitment of epigenetic regulators Sin3, Cyc8 and Tup1
- PMID: 32980916
- PMCID: PMC7599196
- DOI: 10.1007/s00294-020-01109-4
Functional analysis of Cti6 core domain responsible for recruitment of epigenetic regulators Sin3, Cyc8 and Tup1
Abstract
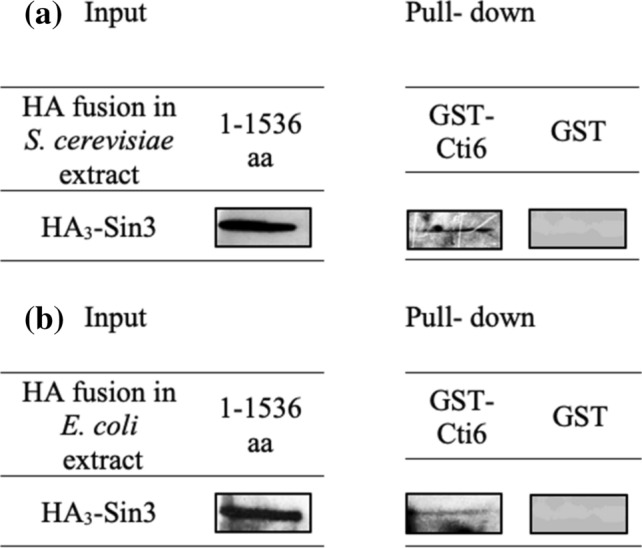
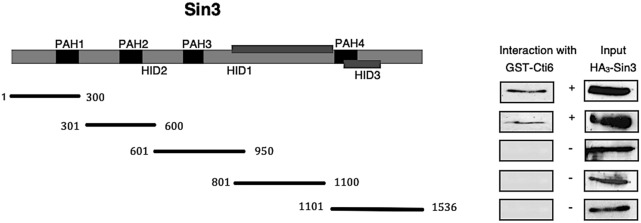
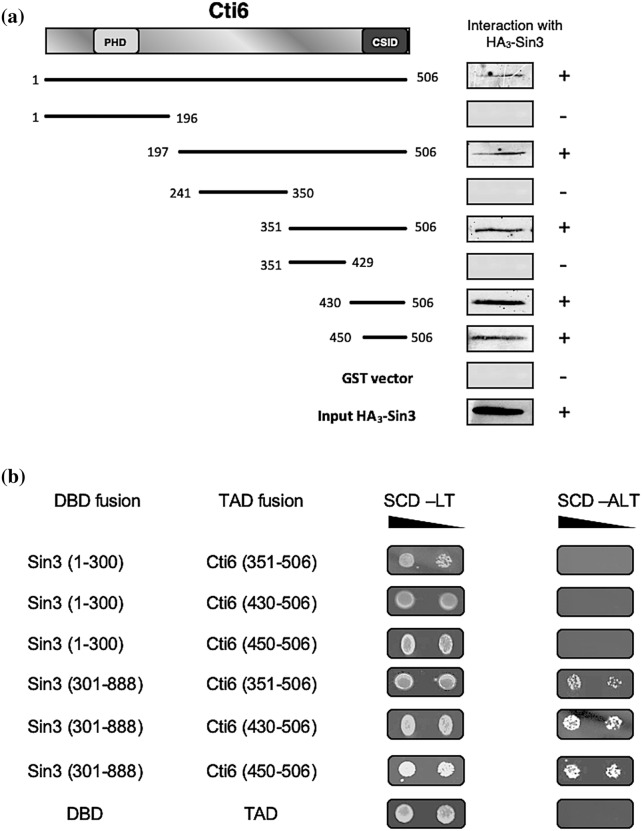
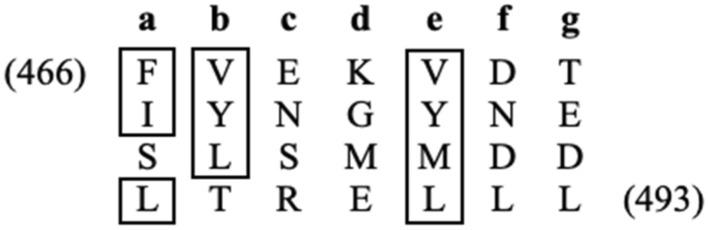
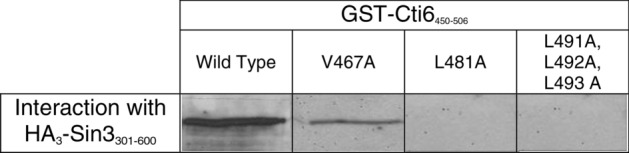
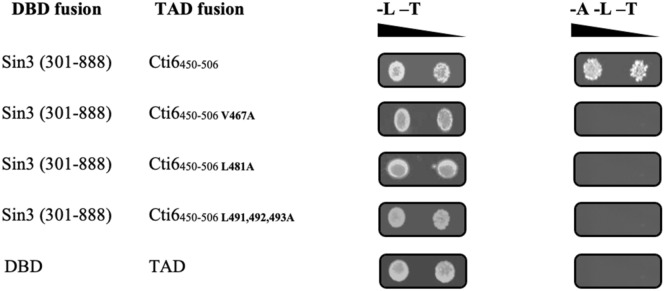
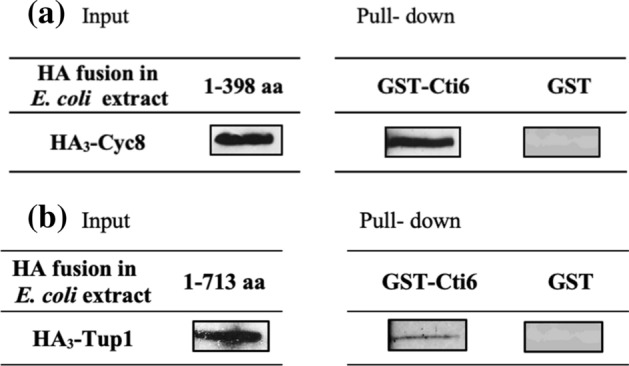
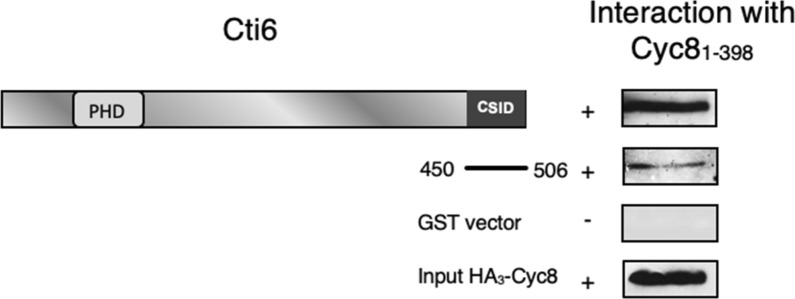
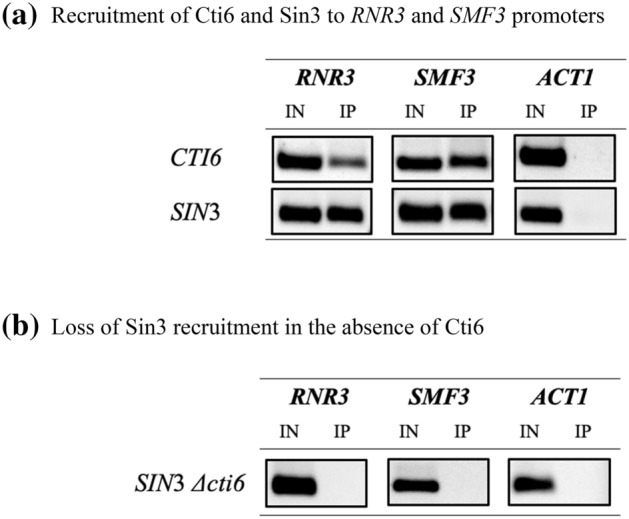
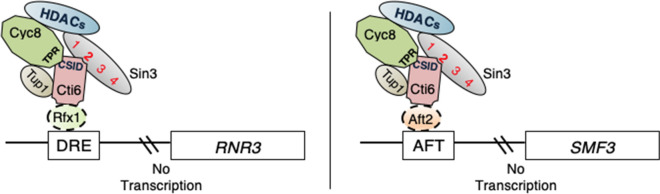
Mapping of effective protein domains is a demanding stride to disclose the functional relationship between regulatory complexes. Domain analysis of protein interactions is requisite for understanding the pleiotropic responses of the respective partners. Cti6 is a multifunctional regulator for which we could show recruitment of co-repressors Sin3, Cyc8 and Tup1. However, the responsible core domain tethering Cti6 to these co-repressors is poorly understood. Here, we report the pivotal domain of Cti6 that is indispensable for co-repressor recruitment. We substantiated that amino acids 450-506 of Cti6 bind PAH2 of Sin3. To analyse this Cti6-Sin3 Interaction Domain (CSID) in more detail, selected amino acids within CSID were replaced by alanine. It is revealed that hydrophobic amino acids V467, L481 and L491 L492 L493 are important for Cti6-Sin3 binding. In addition to PAH2 of Sin3, CSID also binds to tetratricopeptide repeats (TPR) of Cyc8. Indeed, we could demonstrate Cti6 recruitment to promoters of genes, such as RNR3 and SMF3, containing iron-responsive elements (IRE). Importantly, Sin3 is also recruited to these promoters but only in the presence of functional Cti6. Our findings provide novel insights toward the critical interaction domain in the co-regulator Cti6, which is a component of regulatory complexes that are closely related to chromatin architecture and the epigenetic status of genes that are regulated by pleiotropic co-repressors.
Keywords: Corepressor Sin3; Cti6; Histone deacetylase Rpd3; Protein–protein interaction.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
