

Learned feature variance is encoded in the target template and drives visual search
- PMID: 32982562
- PMCID: PMC7518469
- DOI: 10.1080/13506285.2019.1645779
Learned feature variance is encoded in the target template and drives visual search
Abstract
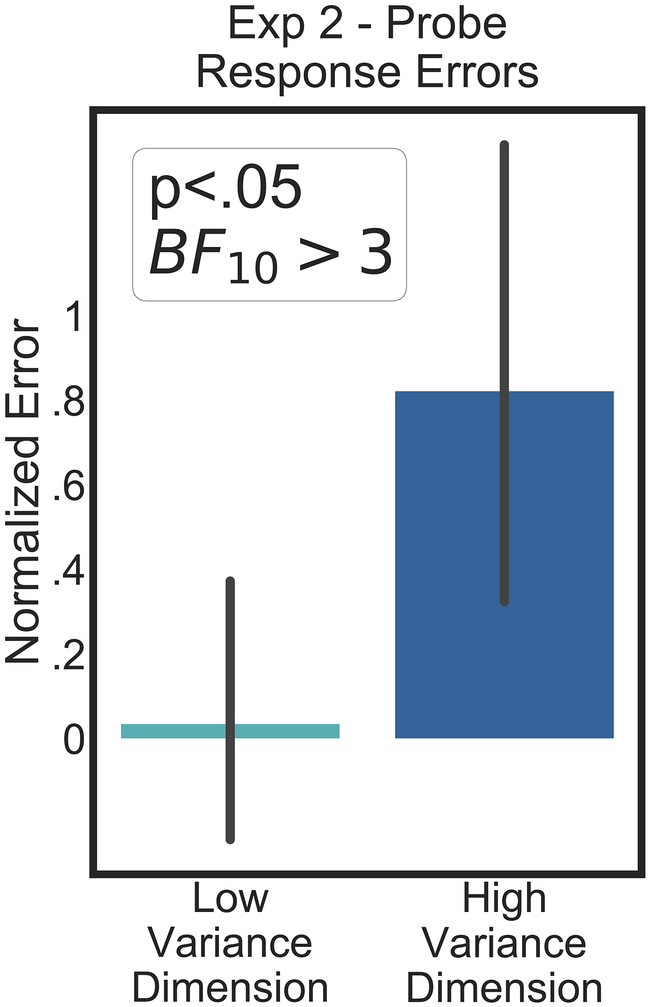
Real world visual search targets are frequently imperfect perceptual matches to our internal target templates. For example, the same friend on different occasions is likely to wear different clothes, hairstyles, and accessories, but some of these may be more likely to vary than others. The ability to deal with template-to-target variability is important to visual search in natural environments, but we know relatively little about how feature variability is handled by the attentional system. In these studies, we test the hypothesis that top-down attentional biases are sensitive to the variance of target feature dimensions over time and prioritize information from less-variable dimensions. On each trial, subjects were shown a target cue composed of colored dots moving in a specific direction followed by a working memory probe (30%) or visual search display (70%). Critically, the target features in the visual search display differed from the cue, with one feature drawn from a distribution narrowly centered over the cued feature (low-variance dimension), and the other sampled from a broader distribution (high-variance dimension). The results demonstrate that subjects used knowledge of the likely cue-to-target variance to set template precision and bias attentional selection. Moreover, an individual's working memory precision for each feature predicted search performance. Our results suggest that observers are sensitive to the variance of feature dimensions within a target and use this information to weight mechanisms of attentional selection.
Keywords: Feature Based Attention; Template; Variability; Visual Attention.
Figures










Similar articles
-
Involuntary top-down control by search-irrelevant features: Visual working memory biases attention in an object-based manner.Cognition. 2018 Mar;172:37-45. doi: 10.1016/j.cognition.2017.12.002. Epub 2017 Dec 8. Cognition. 2018. PMID: 29223864
-
The Role of Color in Search Templates for Real-world Target Objects.J Cogn Neurosci. 2016 Nov;28(11):1714-1727. doi: 10.1162/jocn_a_00996. Epub 2016 Jun 17. J Cogn Neurosci. 2016. PMID: 27315273
-
The precision of attentional selection is far worse than the precision of the underlying memory representation.Cognition. 2019 May;186:20-31. doi: 10.1016/j.cognition.2019.02.001. Epub 2019 Feb 7. Cognition. 2019. PMID: 30739056
-
Activation of new attentional templates for real-world objects in visual search.J Cogn Neurosci. 2015 May;27(5):902-12. doi: 10.1162/jocn_a_00747. Epub 2014 Oct 16. J Cogn Neurosci. 2015. PMID: 25321485
-
Association between cue lead time and template-for-rejection effect.Atten Percept Psychophys. 2019 Aug;81(6):1880-1889. doi: 10.3758/s13414-019-01761-0. Atten Percept Psychophys. 2019. PMID: 31114955 Review.
Cited by
-
Attentional priority is determined by predicted feature distributions.J Exp Psychol Hum Percept Perform. 2022 Nov;48(11):1201-1212. doi: 10.1037/xhp0001041. Epub 2022 Sep 1. J Exp Psychol Hum Percept Perform. 2022. PMID: 36048065 Free PMC article.
-
Task-adaptive changes to the target template in response to distractor context: Separability versus similarity.J Exp Psychol Gen. 2024 Feb;153(2):564-572. doi: 10.1037/xge0001507. Epub 2023 Nov 2. J Exp Psychol Gen. 2024. PMID: 37917441 Free PMC article.
-
Flexible weighting of target features based on distractor context.Atten Percept Psychophys. 2020 Feb;82(2):739-751. doi: 10.3758/s13414-019-01910-5. Atten Percept Psychophys. 2020. PMID: 31741317 Free PMC article. Clinical Trial.
-
Prefrontal Cortex Codes Representations of Target Identity and Feature Uncertainty.J Neurosci. 2023 Dec 13;43(50):8769-8776. doi: 10.1523/JNEUROSCI.1117-23.2023. J Neurosci. 2023. PMID: 37875376 Free PMC article.
-
Attention to object categories: Selection history determines the breadth of attentional tuning during real-world object search.J Exp Psychol Gen. 2024 Jun;153(6):1568-1581. doi: 10.1037/xge0001575. Epub 2024 Apr 22. J Exp Psychol Gen. 2024. PMID: 38647455 Free PMC article.
References
-
- Chelazzi L, Miller EK, Duncan J, & Desimone R (1993). A neural basis for visual search in inferior temporal cortex. Nature, 363(6427), 345–347. Retrieved from http://www.nature.com.globalproxy.cvt.dk/nature/journal/v333/n6176/pdf/3... - PubMed