

Latitudinal Biogeographic Structuring in the Globally Distributed Moss Ceratodon purpureus
- PMID: 32983208
- PMCID: PMC7484499
- DOI: 10.3389/fpls.2020.502359
Latitudinal Biogeographic Structuring in the Globally Distributed Moss Ceratodon purpureus
Abstract
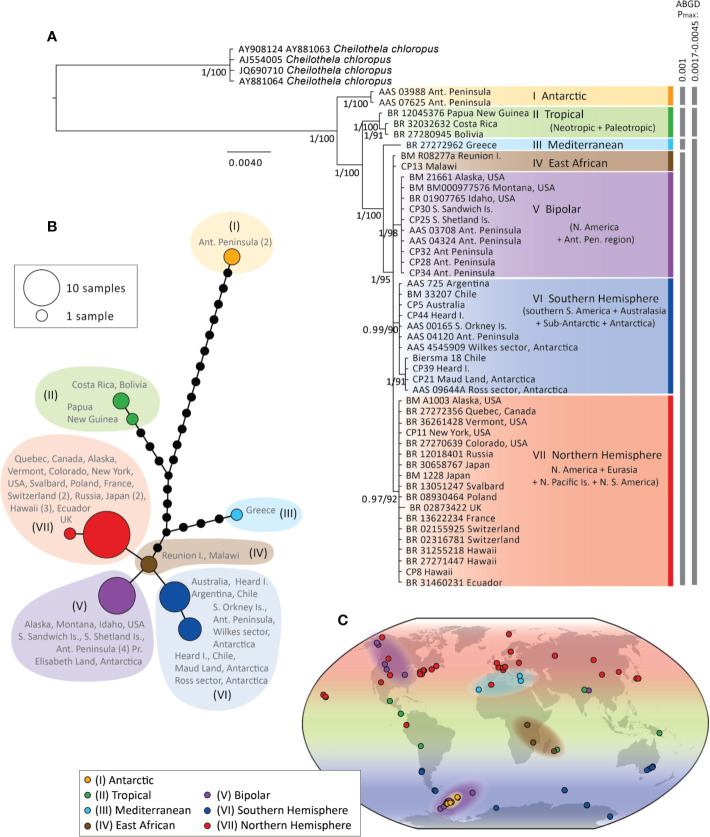
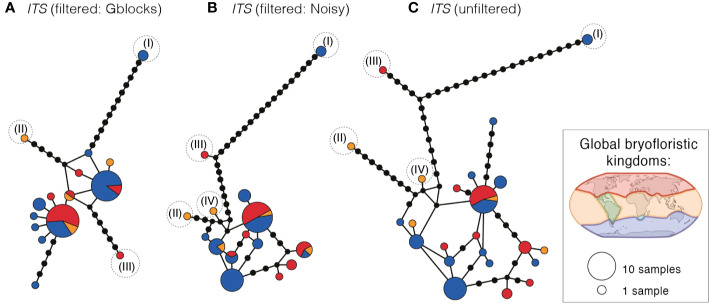
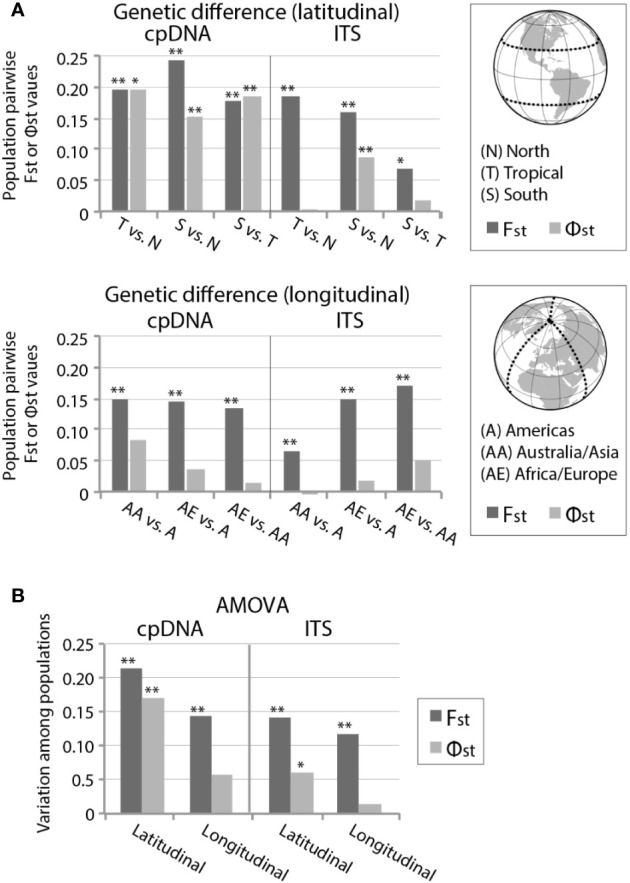
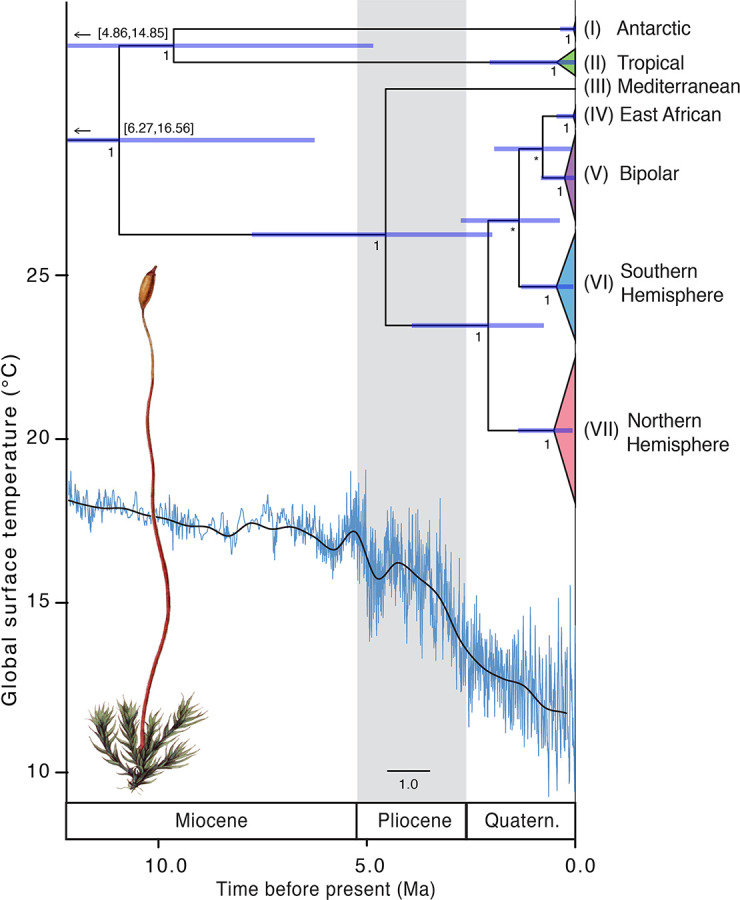
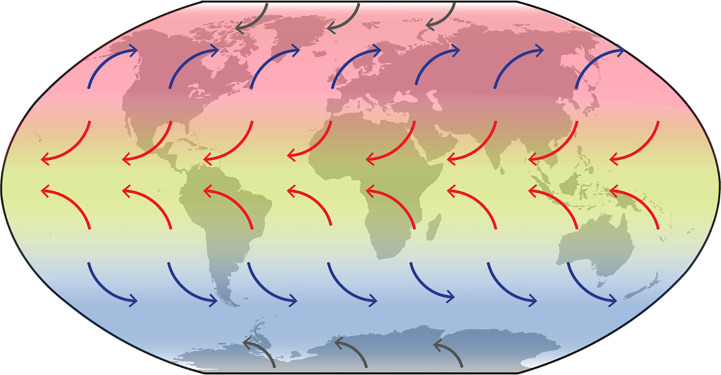
Biogeographic patterns of globally widespread species are expected to reflect regional structure, as well as connectivity caused by occasional long-distance dispersal. We assessed the level and drivers of population structure, connectivity, and timescales of population isolation in one of the most widespread and ruderal plants in the world - the common moss Ceratodon purpureus. We applied phylogenetic, population genetic, and molecular dating analyses to a global (n = 147) sampling data set, using three chloroplast loci and one nuclear locus. The plastid data revealed several distinct and geographically structured lineages, with connectivity patterns associated with worldwide, latitudinal "bands." These imply that connectivity is strongly influenced by global atmospheric circulation patterns, with dispersal and establishment beyond these latitudinal bands less common. Biogeographic patterns were less clear within the nuclear marker, with gene duplication likely hindering the detection of these. Divergence time analyses indicated that the current matrilineal population structure in C. purpureus has developed over the past six million years, with lineages diverging during the late Miocene, Pliocene, and Quaternary. Several colonization events in the Antarctic were apparent, as well as one old and distinct Antarctic clade, possibly isolated on the continent since the Pliocene. As C. purpureus is considered a model organism, the matrilineal biogeographic structure identified here provides a useful framework for future genetic and developmental studies on bryophytes. Our general findings may also be relevant to understanding global environmental influences on the biogeography of other organisms with microscopic propagules (e.g., spores) dispersed by wind.
Keywords: Antarctica; bryophyte; global; model organism; moss; phylogeography; spore; wind.
Copyright © 2020 Biersma, Convey, Wyber, Robinson, Dowton, van de Vijver, Linse, Griffiths and Jackson.
Figures
References
-
- Biersma E. M., Jackson J. A., Bracegirdle T. J., Griffiths H., Linse K., Convey P. (2018. a). Low genetic variation between South American and Antarctic populations of the bank−forming moss Chorisodontium aciphyllum (Dicranaceae). Polar. Biol. 41, 599–610. 10.1007/s00300-017-2221-1 - DOI
-
- Biersma E. M., Jackson J. A., Stech M., Griffiths H., Linse K., Convey P. (2018. b). Molecular data suggest long-term in situ Antarctic persistence within Antarctica’s most speciose plant genus, Schistidium. Front. Ecol. Evol. 6:77. 10.3389/fevo.2018.00077 - DOI
-
- Biersma E. M., Torres-Díaz C., Molina-Montenegro M. A., Newsham K. K., Vidal M. A., Collado G. A., et al. (2020). Multiple post-glacial colonisation events of the Antarctic pearlwort Colobanthus quitensis (Caryophyllaceae) reveal the recent arrival of native Antarctic vascular flora. J. Biogeogr. 00, 1–11. 10.1111/jbi.13843 - DOI
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous