

Forces to Drive Neuronal Migration Steps
- PMID: 32984342
- PMCID: PMC7490296
- DOI: 10.3389/fcell.2020.00863
Forces to Drive Neuronal Migration Steps
Abstract
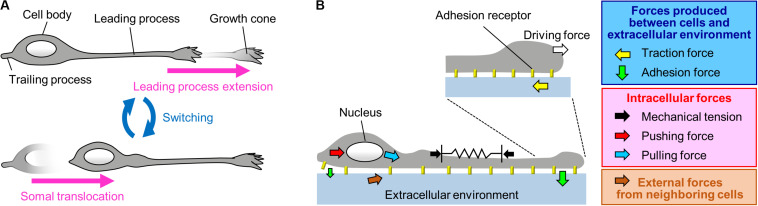
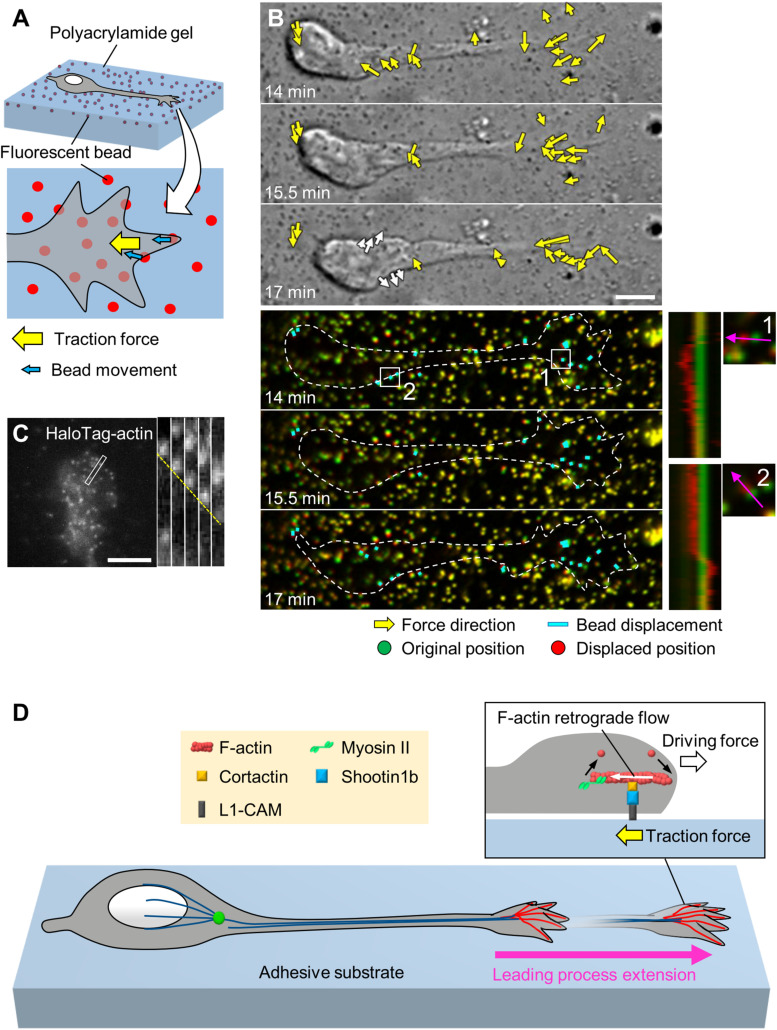
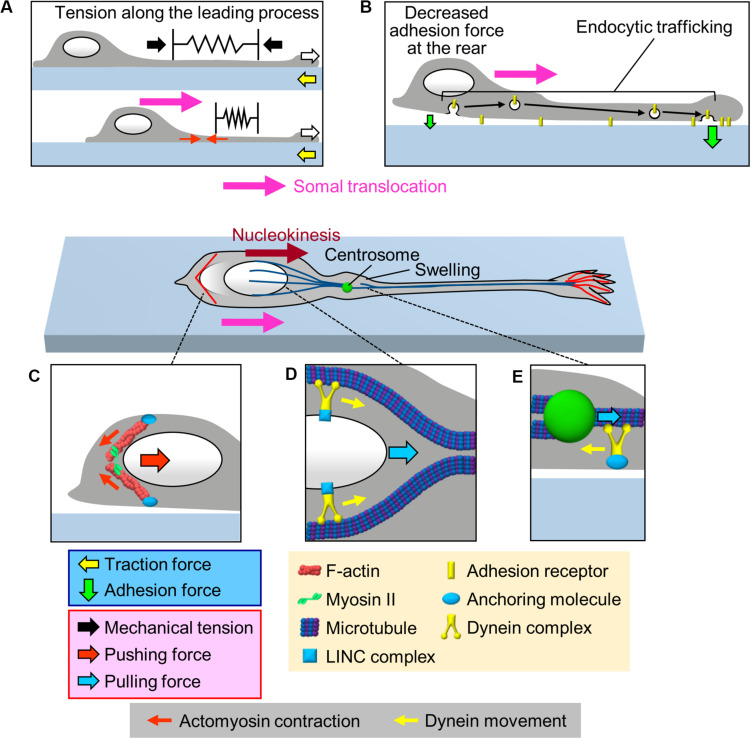
To establish and maintain proper brain architecture and elaborate neural networks, neurons undergo massive migration. As a unique feature of their migration, neurons move in a saltatory manner by repeating two distinct steps: extension of the leading process and translocation of the cell body. Neurons must therefore generate forces to extend the leading process as well as to translocate the cell body. In addition, neurons need to switch these forces alternately in order to orchestrate their saltatory movement. Recent studies with mechanobiological analyses, including traction force microscopy, cell detachment analyses, live-cell imaging, and loss-of-function analyses, have begun to reveal the forces required for these steps and the molecular mechanics underlying them. Spatiotemporally organized forces produced between cells and their extracellular environment, as well as forces produced within cells, play pivotal roles to drive these neuronal migration steps. Traction force produced by the leading process growth cone extends the leading processes. On the other hand, mechanical tension of the leading process, together with reduction in the adhesion force at the rear and the forces to drive nucleokinesis, translocates the cell body. Traction forces are generated by mechanical coupling between actin filament retrograde flow and the extracellular environment through clutch and adhesion molecules. Forces generated by actomyosin and dynein contribute to the nucleokinesis. In addition to the forces generated in cell-intrinsic manners, external forces provided by neighboring migratory cells coordinate cell movement during collective migration. Here, we review our current understanding of the forces that drive neuronal migration steps and describe the molecular machineries that generate these forces for neuronal migration.
Keywords: actomyosin; adhesion force; dynein; mechanical tension; mechanobiology; neuronal migration; shootin1; traction force.
Copyright © 2020 Minegishi and Inagaki.
Figures
References
-
- Bellion A., Baudoin J. P., Alvarez C., Bornens M., Metin C. (2005). Nucleokinesis in tangentially migrating neurons comprises two alternating phases: forward migration of the Golgi/centrosome associated with centrosome splitting and myosin contraction at the rear. J. Neurosci. 25 5691–5699. 10.1523/JNEUROSCI.1030-05.2005 - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Research Materials