

Identification of tannic cell walls at the outer surface of the endosperm upon Arabidopsis seed coat rupture
- PMID: 32985026
- PMCID: PMC7702108
- DOI: 10.1111/tpj.14994
Identification of tannic cell walls at the outer surface of the endosperm upon Arabidopsis seed coat rupture
Abstract
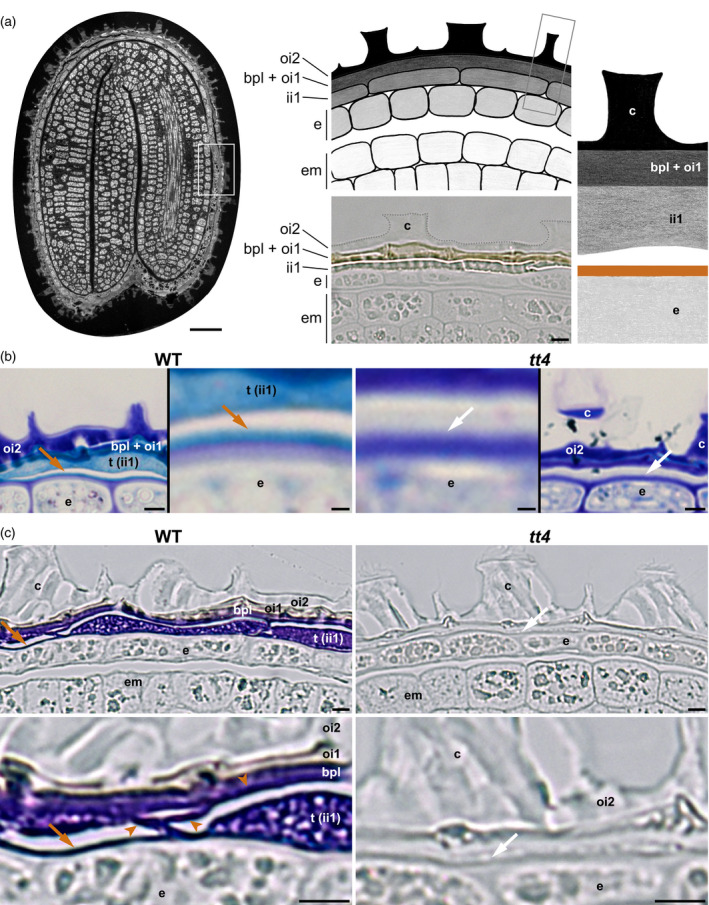
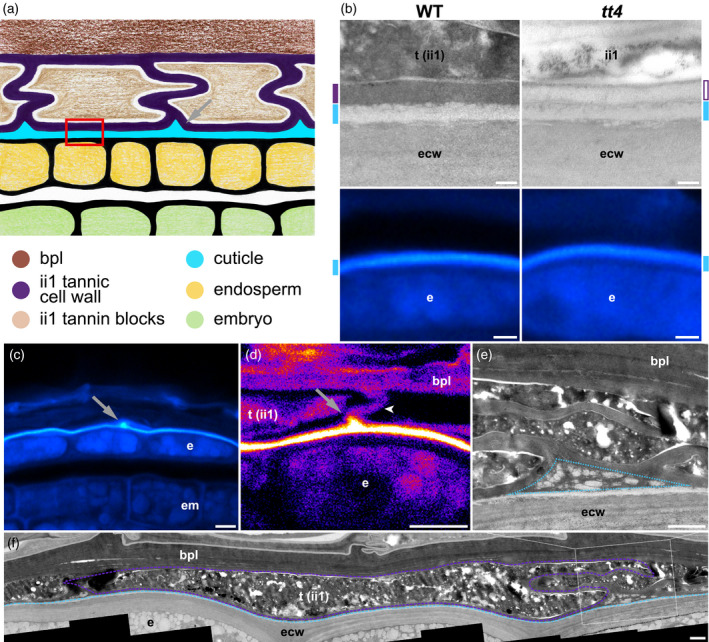
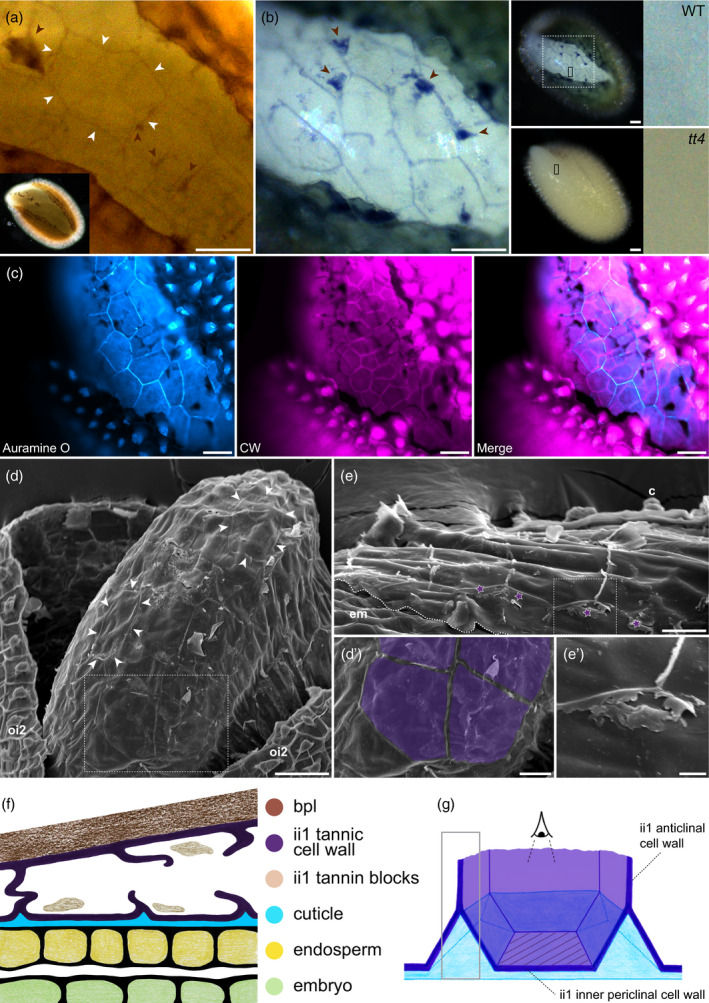
The seed coat is specialized dead tissue protecting the plant embryo from mechanical and oxidative damage. Tannins, a type of flavonoids, are antioxidants known to accumulate in the Arabidopsis seed coat and transparent testa mutant seeds, deficient in flavonoid synthesis, exhibit low viability. However, their precise contribution to seed coat architecture and biophysics remains evasive. A seed coat cuticle, covering the endosperm outer surface and arising from the seed coat inner integument 1 cell layer was, intriguingly, previously shown to be more permeable in transparent testa mutants deficient not in cuticular component synthesis, but rather in flavonoid synthesis. Investigating the role of flavonoids in cuticle permeability led us to identify periclinal inner integument 1 tannic cell walls being attached, together with the cuticle, to the endosperm surface upon seed coat rupture. Hence, inner integument 1 tannic cell walls and the cuticle form two fused layers present at the surface of the exposed endosperm upon seed coat rupture, regulating its permeability. Their potential physiological role during seed germination is discussed.
Keywords: Arabidopsis thaliana; apoplastic barrier; endosperm; seed; seed coat; tannic cell wall.
© 2020 The Authors. The Plant Journal published by Society for Experimental Biology and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Comment in
-
Secrets of the seed: uncovering the real identity of the endosperm-surrounding cuticle.Plant J. 2020 Nov;104(3):565-566. doi: 10.1111/tpj.15008. Plant J. 2020. PMID: 33118278 No abstract available.
References
-
- Ancillotto, L. , Sozio, G. and Mortelliti, A. (2015) Acorns were good until tannins were found: Factors affecting seed‐selection in the hazel dormouse (Muscardinus avellanarius). Mamm Biol. 80, 135–140.
-
- Andème Ondzighi, C. , Christopher, D.A. , Cho, E.J. , Chang, S.‐C. and Staehelin, L.A. (2008) Arabidopsis protein disulfide isomerase‐5 inhibits cysteine proteases during trafficking to vacuoles before programmed cell death of the endothelium in developing seeds. Plant Cell, 20, 2205–2220. - PMC - PubMed
-
- Auger, B. , Marnet, N. , Gautier, V. , Maia‐Grondard, A. , Leprince, F. , Renard, M. , Guyot, S. , Nesi, N. and Routaboul, J.‐M. (2010) A Detailed survey of seed coat flavonoids in developing seeds of Brassica napus L. J. Agric. Food Chem. 58, 6246–6256. - PubMed
-
- Beeckman, T. , De Rycke, R. , Viane, R. and Inzé, D. (2000) Histological study of seed coat development in Arabidopsis thaliana. J. Plant Res. 113, 139–148.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
