

Cocoonase is indispensable for Lepidoptera insects breaking the sealed cocoon
- PMID: 32986696
- PMCID: PMC7544147
- DOI: 10.1371/journal.pgen.1009004
Cocoonase is indispensable for Lepidoptera insects breaking the sealed cocoon
Abstract
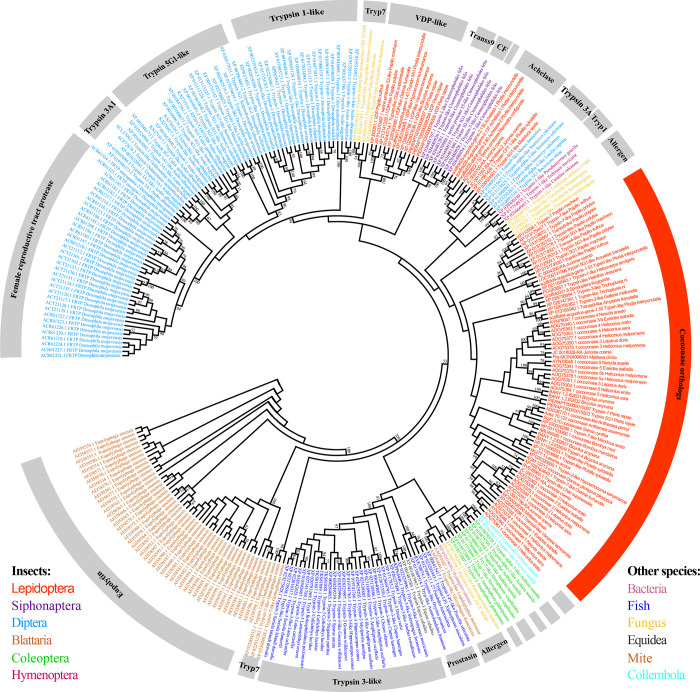
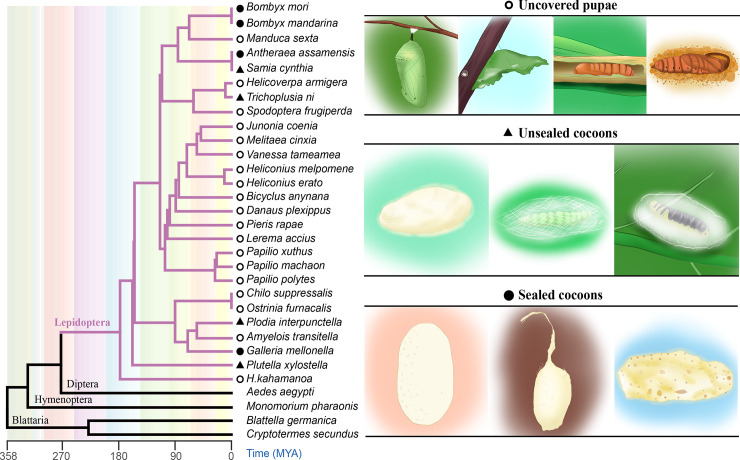
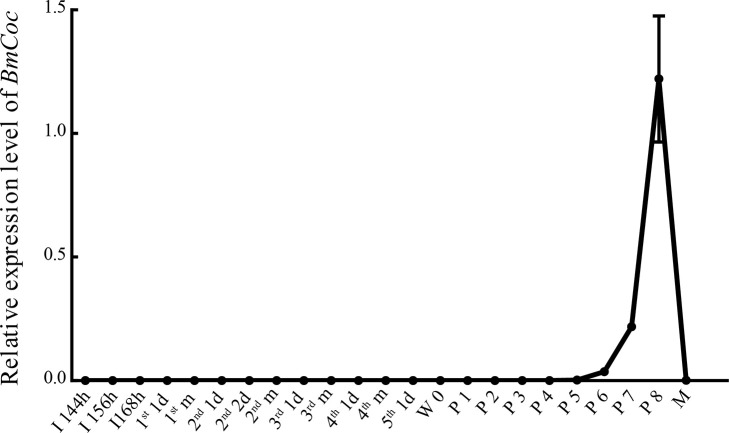
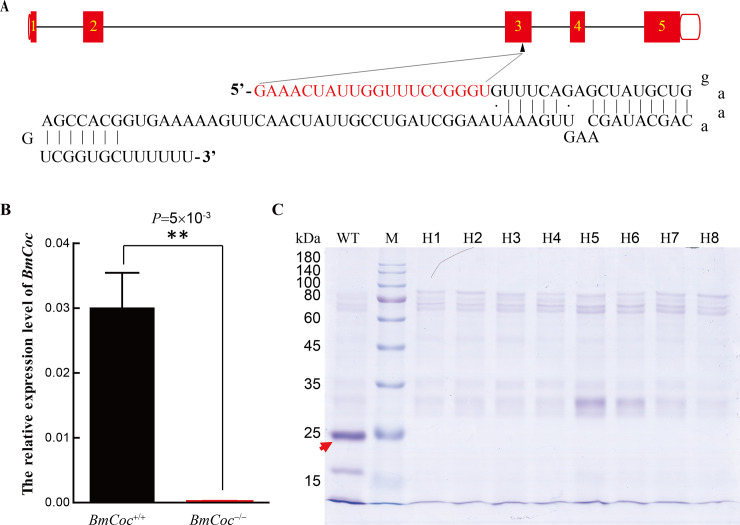
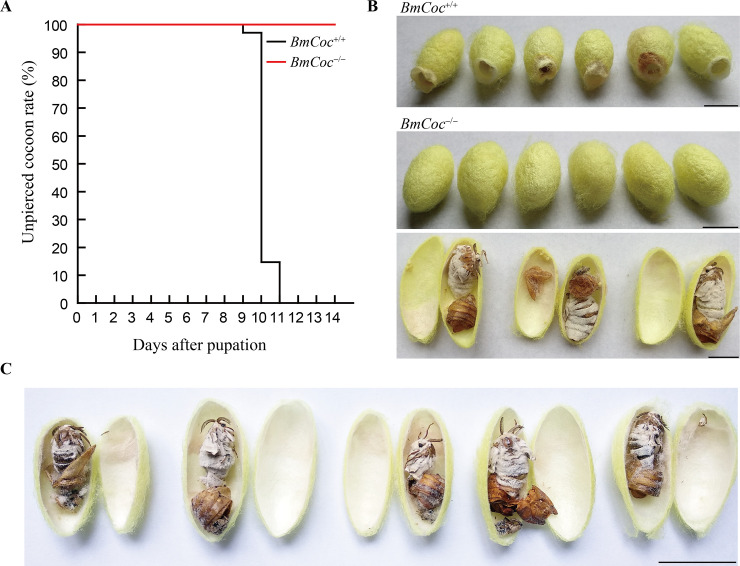
Many insects spin cocoons to protect the pupae from unfavorable environments and predators. After emerging from the pupa, the moths must escape from the sealed cocoons. Previous works identified cocoonase as the active enzyme loosening the cocoon to form an escape-hatch. Here, using bioinformatics tools, we show that cocoonase is specific to Lepidoptera and that it probably existed before the occurrence of lepidopteran insects spinning cocoons. Despite differences in cocooning behavior, we further show that cocoonase evolved by purification selection in Lepidoptera and that the selection is more intense in lepidopteran insects spinning sealed cocoons. Experimentally, we applied gene editing techniques to the silkworm Bombyx mori, which spins a dense and sealed cocoon, as a model of lepidopteran insects spinning sealed cocoons. We knocked out cocoonase using the CRISPR/Cas9 system. The adults of homozygous knock-out mutants were completely formed and viable but stayed trapped and died naturally in the cocoon. This is the first experimental and phenotypic evidence that cocoonase is the determining factor for breaking the cocoon. This work led to a novel silkworm strain yielding permanently intact cocoons and provides a new strategy for controlling the pests that form cocoons.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Ruxton GD, Sherratt TN, Speed MP. Avoiding attack: the evolutionary ecology of crypsis, warning signals and mimicry. Oxford University Press, Oxford, UK; 2004.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
