

Structural basis of DNA replication origin recognition by human Orc6 protein binding with DNA
- PMID: 32986843
- PMCID: PMC7641730
- DOI: 10.1093/nar/gkaa751
Structural basis of DNA replication origin recognition by human Orc6 protein binding with DNA
Abstract
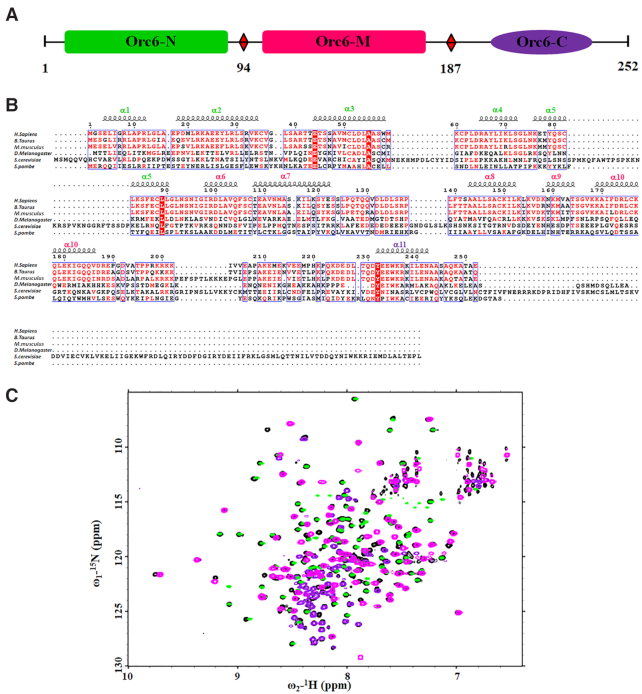
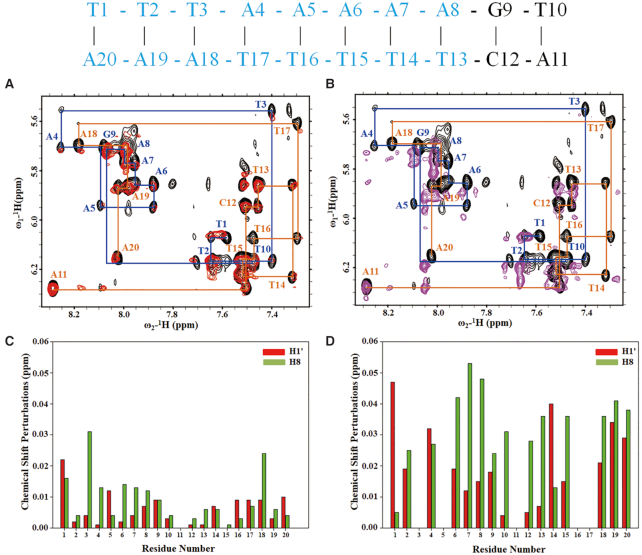
The six-subunit origin recognition complex (ORC), a DNA replication initiator, defines the localization of the origins of replication in eukaryotes. The Orc6 subunit is the smallest and the least conserved among ORC subunits. It is required for DNA replication and essential for viability in all species. Orc6 in metazoans carries a structural homology with transcription factor TFIIB and can bind DNA on its own. Here, we report a solution structure of the full-length human Orc6 (HsOrc6) alone and in a complex with DNA. We further showed that human Orc6 is composed of three independent domains: N-terminal, middle and C-terminal (HsOrc6-N, HsOrc6-M and HsOrc6-C). We also identified a distinct DNA-binding domain of human Orc6, named as HsOrc6-DBD. The detailed analysis of the structure revealed novel amino acid clusters important for the interaction with DNA. Alterations of these amino acids abolish DNA-binding ability of Orc6 and result in reduced levels of DNA replication. We propose that Orc6 is a DNA-binding subunit of human/metazoan ORC and may play roles in targeting, positioning and assembling the functional ORC at the origins.
© The Author(s) 2020. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
