

Reversal of diet-induced hepatic steatosis by peripheral CB1 receptor blockade in mice is p53/miRNA-22/SIRT1/PPARα dependent
- PMID: 32987186
- PMCID: PMC7563015
- DOI: 10.1016/j.molmet.2020.101087
Reversal of diet-induced hepatic steatosis by peripheral CB1 receptor blockade in mice is p53/miRNA-22/SIRT1/PPARα dependent
Abstract
Objective: The endocannabinoid (eCB) system is increasingly recognized as being crucially important in obesity-related hepatic steatosis. By activating the hepatic cannabinoid-1 receptor (CB1R), eCBs modulate lipogenesis and fatty acid oxidation. However, the underlying molecular mechanisms are largely unknown.
Methods: We combined unbiased bioinformatics techniques, mouse genetic manipulations, multiple pharmacological, molecular, and cellular biology approaches, and genomic sequencing to systematically decipher the role of the hepatic CB1R in modulating fat utilization in the liver and explored the downstream molecular mechanisms.
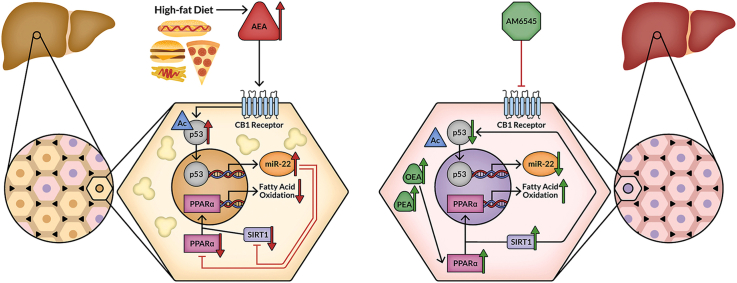
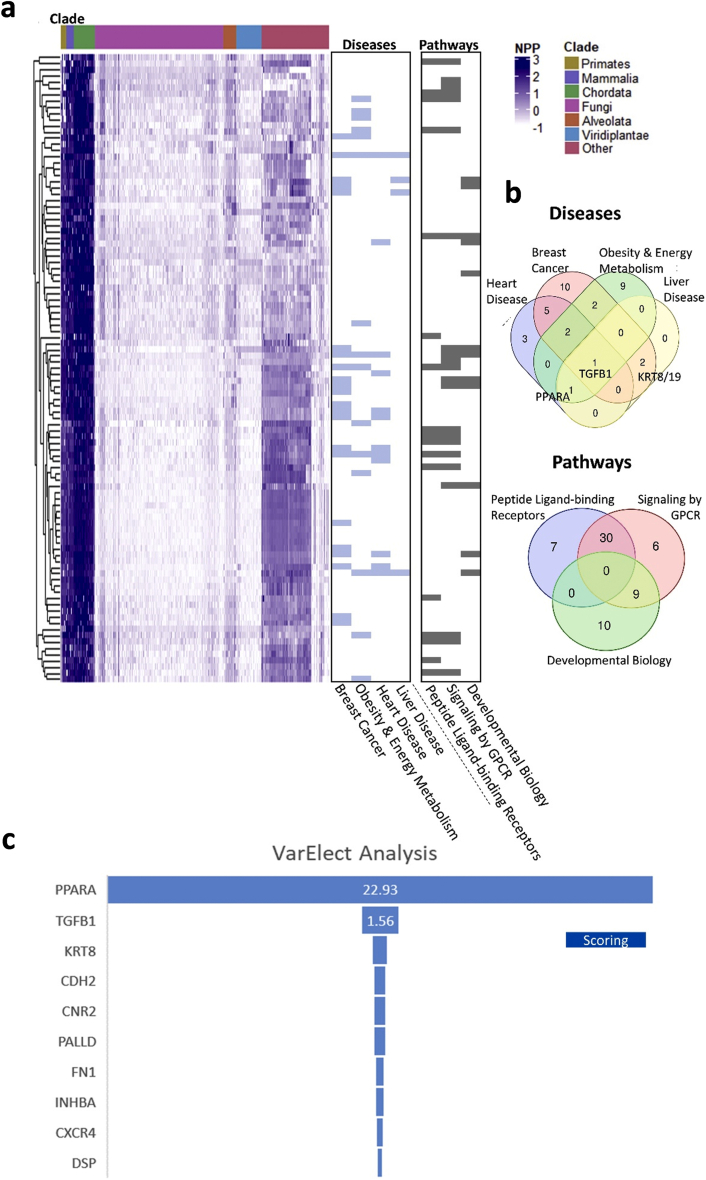
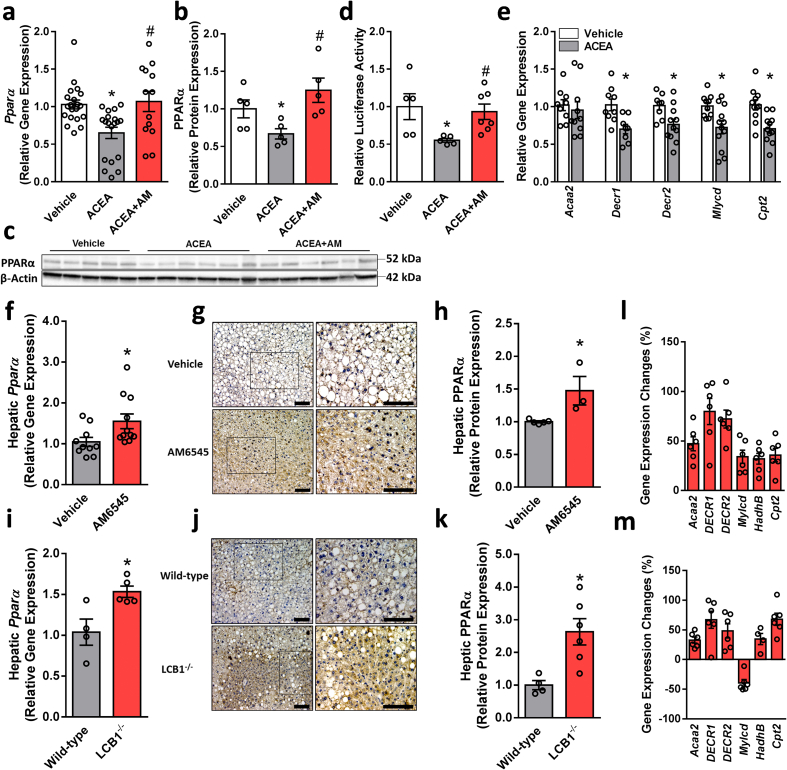
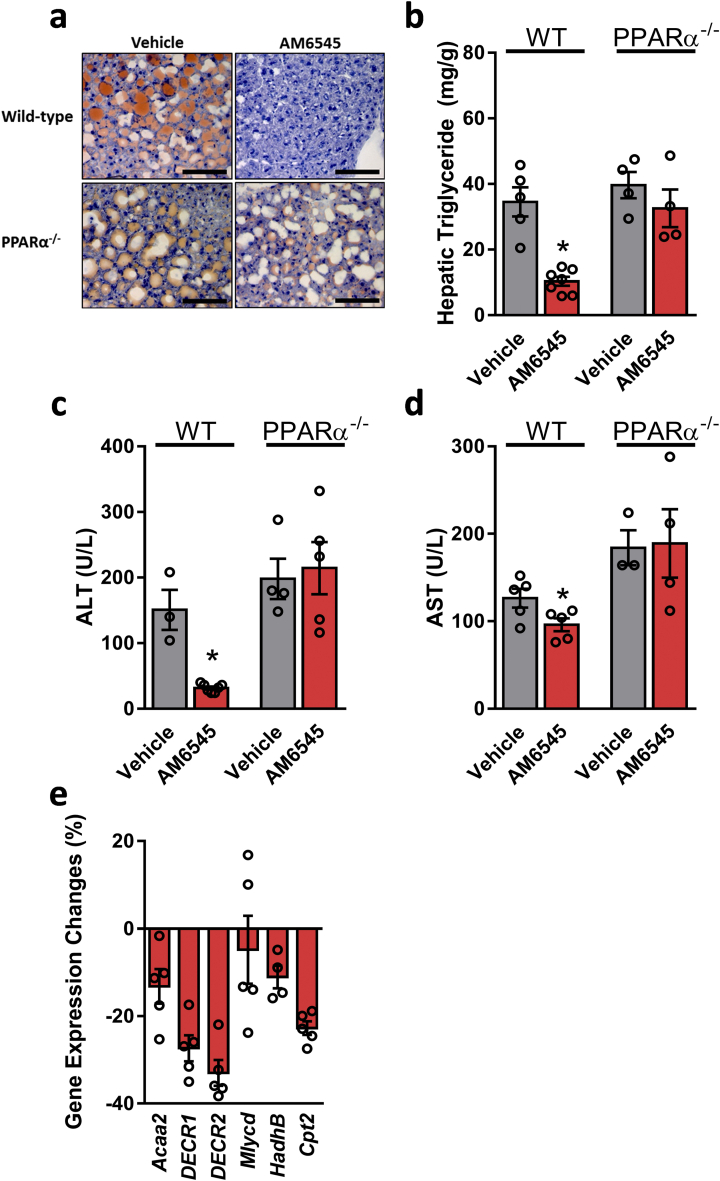
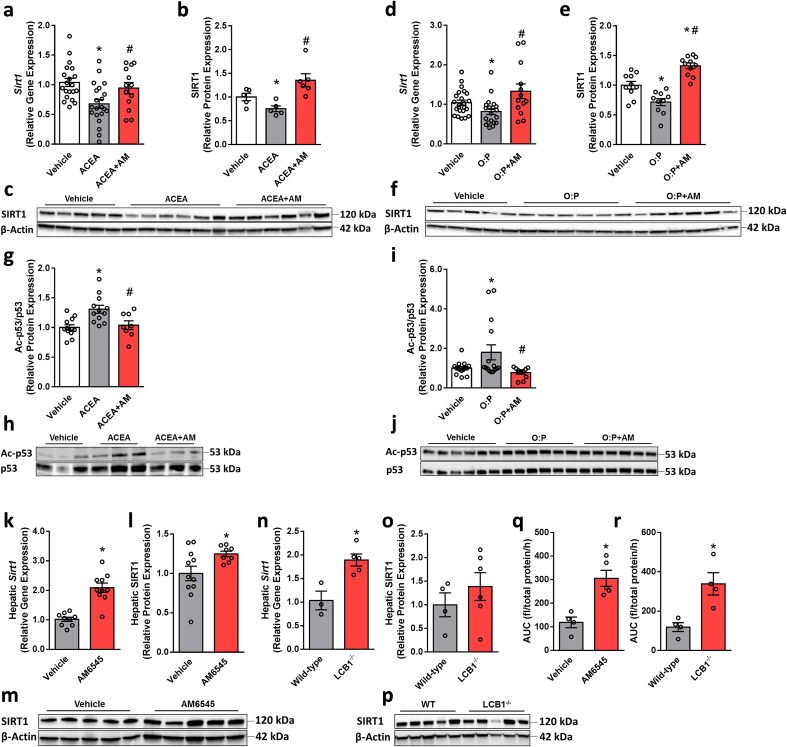
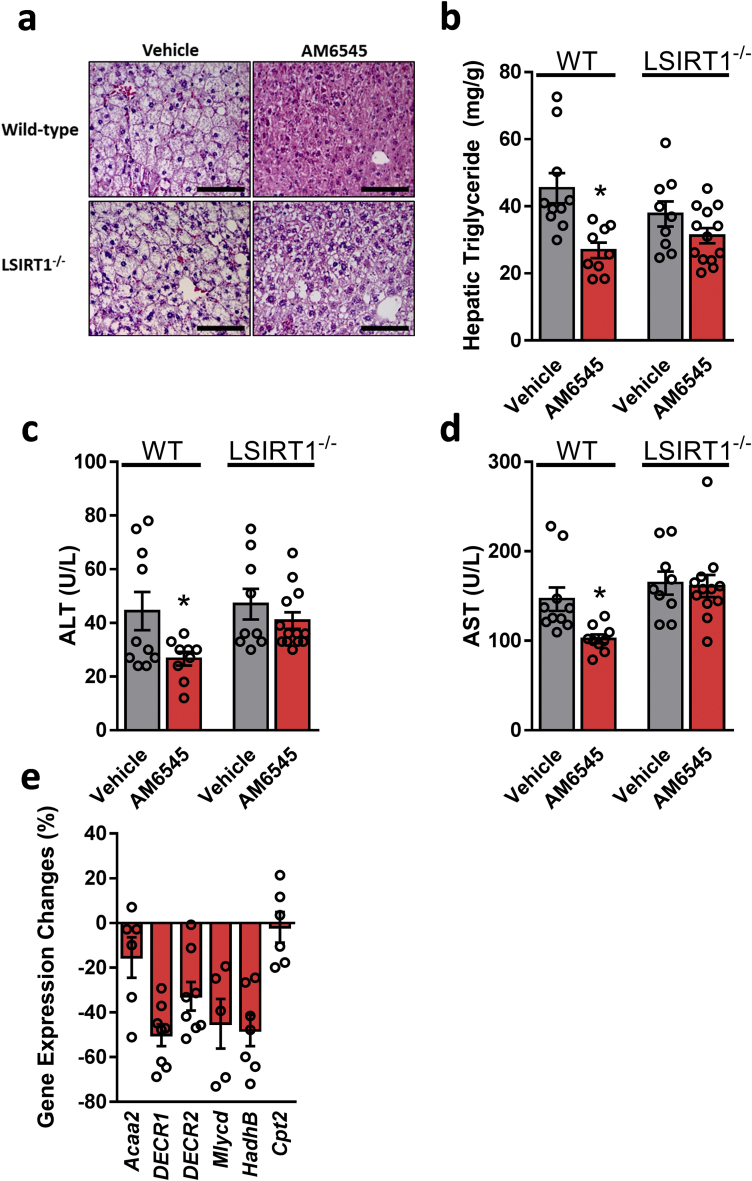
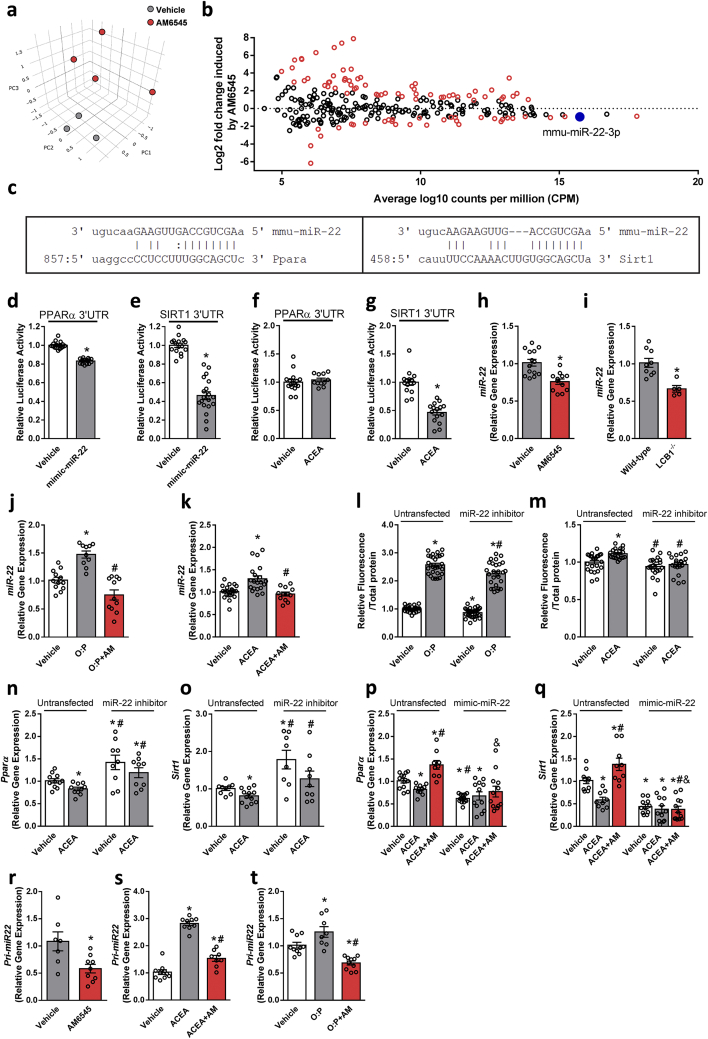
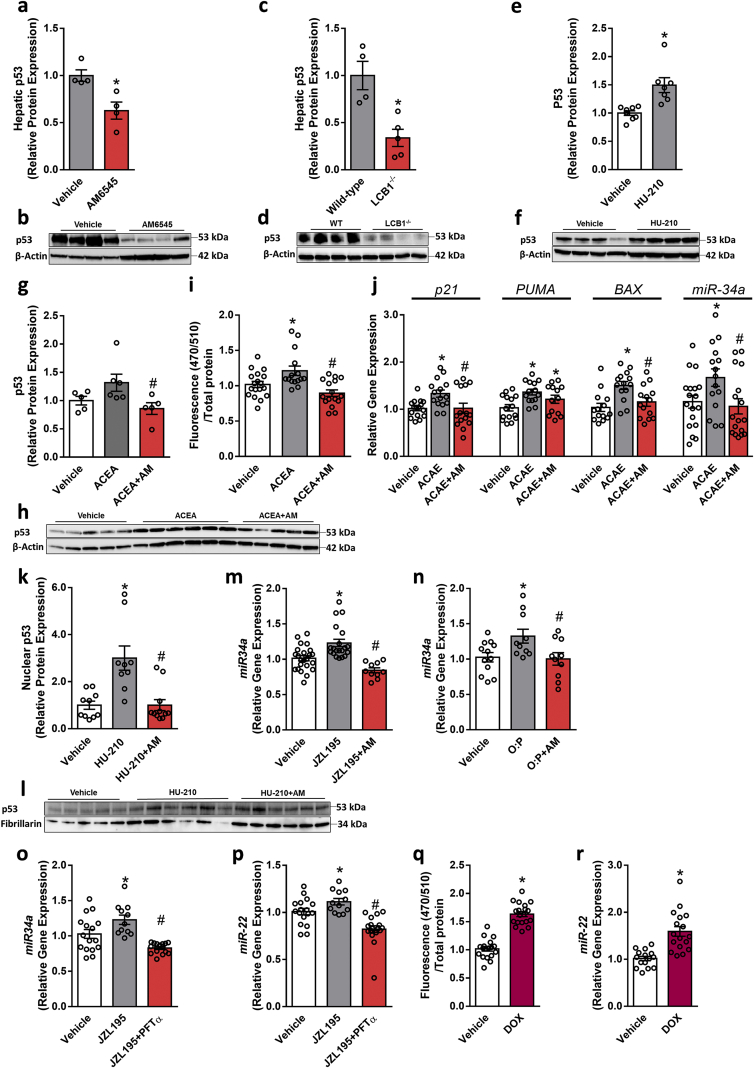
Results: Using an unbiased normalized phylogenetic profiling analysis, we found that the CB1R evolutionarily coevolves with peroxisome proliferator-activated receptor-alpha (PPARα), a key regulator of hepatic lipid metabolism. In diet-induced obese (DIO) mice, peripheral CB1R blockade (using AM6545) induced the reversal of hepatic steatosis and improved liver injury in WT, but not in PPARα-/- mice. The antisteatotic effect mediated by AM6545 in WT DIO mice was accompanied by increased hepatic expression and activity of PPARα as well as elevated hepatic levels of the PPARα-activating eCB-like molecules oleoylethanolamide and palmitoylethanolamide. Moreover, AM6545 was unable to rescue hepatic steatosis in DIO mice lacking liver sirtuin 1 (SIRT1), an upstream regulator of PPARα. Both of these signaling molecules were modulated by the CB1R as measured in hepatocytes exposed to lipotoxic conditions or treated with CB1R agonists in the absence/presence of AM6545. Furthermore, using microRNA transcriptomic profiling, we found that the CB1R regulated the hepatic expression, acetylation, and transcriptional activity of p53, resulting in the enhanced expression of miR-22, which was found to specifically target SIRT1 and PPARα.
Conclusions: We provide strong evidence for a functional role of the p53/miR-22/SIRT1/PPARα signaling pathway in potentially mediating the antisteatotic effect of peripherally restricted CB1R blockade.
Keywords: Endocannabinoids; Fatty liver; Nuclear receptor; Obesity; microRNAs.
Copyright © 2020 The Authors. Published by Elsevier GmbH.. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
