

Multiple genetic programs contribute to CD4 T cell memory differentiation and longevity by maintaining T cell quiescence
- PMID: 32987276
- PMCID: PMC7737224
- DOI: 10.1016/j.cellimm.2020.104210
Multiple genetic programs contribute to CD4 T cell memory differentiation and longevity by maintaining T cell quiescence
Abstract
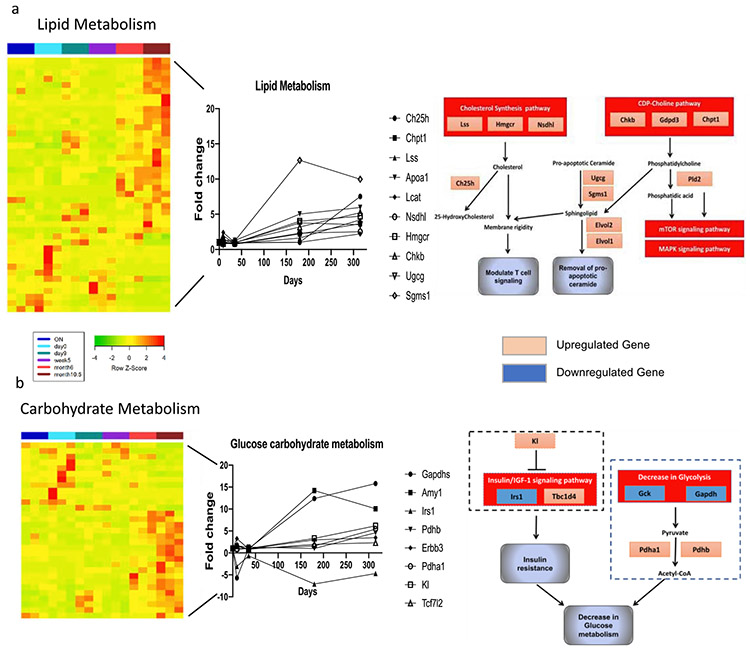
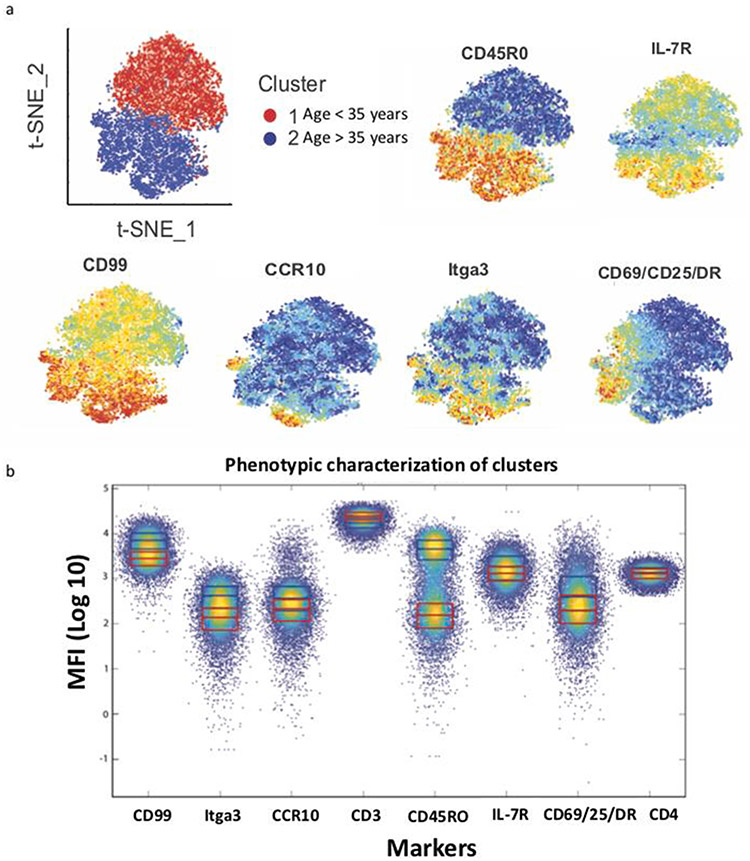
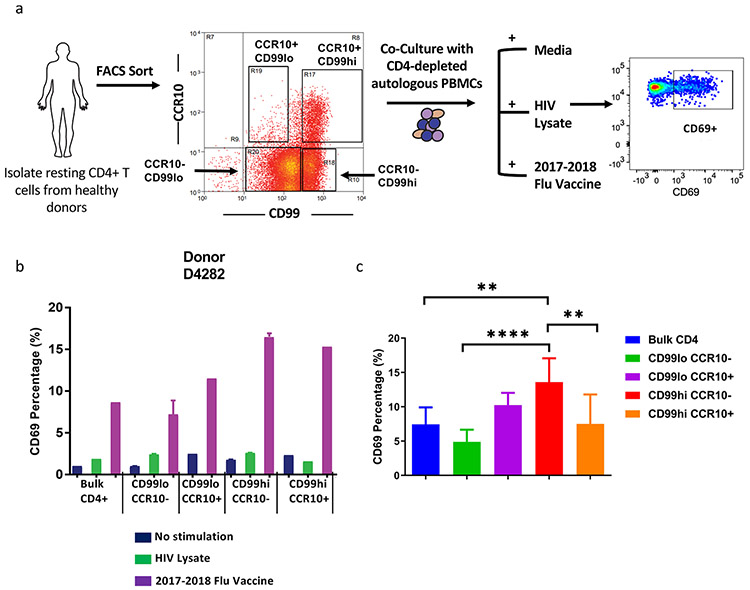
While memory T-cells represent a hallmark of adaptive immunity, little is known about the genetic mechanisms regulating the longevity of memory CD4 T cells. Here, we studied the dynamics of gene expression in antigen specific CD4 T cells during infection, memory differentiation, and long-term survival up to nearly a year in mice. We observed that differentiation into long lived memory cells is associated with increased expression of genes inhibiting cell proliferation and apoptosis as well as genes promoting DNA repair response, lipid metabolism, and insulin resistance. We identified several transmembrane proteins in long-lived murine memory CD4 T cells, which co-localized exclusively within the responding antigen-specific memory CD4 T cells in human. The unique gene signatures of long-lived memory CD4 T cells, along with the new markers that we have defined, will enable a deeper understanding of memory CD4 T cell biology and allow for designing novel vaccines and therapeutics.
Keywords: CD4 T cell; Cell longevity; Gene; Genetic programs; Memory T cell; Memory cell markers.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures




Similar articles
-
Rapid culling of the CD4+ T cell repertoire in the transition from effector to memory.Immunity. 2008 Apr;28(4):533-45. doi: 10.1016/j.immuni.2008.02.014. Epub 2008 Mar 20. Immunity. 2008. PMID: 18356084 Free PMC article.
-
Bcl6 is essential for the generation of long-term memory CD4+ T cells.Int Immunol. 2007 Apr;19(4):427-33. doi: 10.1093/intimm/dxm007. Epub 2007 Feb 16. Int Immunol. 2007. PMID: 17307796
-
Regulatory dendritic cells program generation of interleukin-4-producing alternative memory CD4 T cells with suppressive activity.Blood. 2011 Jan 27;117(4):1218-27. doi: 10.1182/blood-2010-05-285494. Epub 2010 Nov 4. Blood. 2011. PMID: 21051555
-
How Does B Cell Antigen Presentation Affect Memory CD4 T Cell Differentiation and Longevity?Front Immunol. 2021 Jun 10;12:677036. doi: 10.3389/fimmu.2021.677036. eCollection 2021. Front Immunol. 2021. PMID: 34177919 Free PMC article. Review.
-
Similarities and differences in CD4+ and CD8+ effector and memory T cell generation.Nat Immunol. 2003 Sep;4(9):835-42. doi: 10.1038/ni969. Nat Immunol. 2003. PMID: 12942084 Review.
Cited by
-
Advances and challenges in identifying precursors of memory CD4+ T cells.Front Immunol. 2025 May 5;16:1540045. doi: 10.3389/fimmu.2025.1540045. eCollection 2025. Front Immunol. 2025. PMID: 40391228 Free PMC article. Review.
-
Immune Enhancement of Nanoparticle-Encapsulated Ginseng Stem-Leaf Saponins on Porcine Epidemic Diarrhea Virus Vaccine in Mice.Vaccines (Basel). 2022 Oct 27;10(11):1810. doi: 10.3390/vaccines10111810. Vaccines (Basel). 2022. PMID: 36366319 Free PMC article.
-
The Differentiation and Maintenance of SARS-CoV-2-Specific Follicular Helper T Cells.Front Cell Infect Microbiol. 2022 Jul 14;12:953022. doi: 10.3389/fcimb.2022.953022. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35909969 Free PMC article. Review.
-
How a Proposed Hypothesis during My PhD Training Shaped My Career.Crit Rev Immunol. 2020;40(5):449-464. doi: 10.1615/CritRevImmunol.2020035324. Crit Rev Immunol. 2020. PMID: 33463956 Free PMC article.
-
A Novel Immune-Gene Pair Signature Revealing the Tumor Microenvironment Features and Immunotherapy Prognosis of Muscle-Invasive Bladder Cancer.Front Genet. 2021 Nov 26;12:764184. doi: 10.3389/fgene.2021.764184. eCollection 2021. Front Genet. 2021. PMID: 34899849 Free PMC article.
References
-
- Appay V, Bosio A, Lokan S, Wiencek Y, Biervert C, Kusters D, Devevre E, Speiser D, Romero P, Rufer N, Leyvraz S, Sensitive gene expression profiling of human T cell subsets reveals parallel post-thymic differentiation for CD4 + and CD8+ lineages, J Immunol 179 (2007) 7406–7414. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
