

The molecular structure of mammalian primary cilia revealed by cryo-electron tomography
- PMID: 32989303
- PMCID: PMC7610599
- DOI: 10.1038/s41594-020-0507-4
The molecular structure of mammalian primary cilia revealed by cryo-electron tomography
Abstract
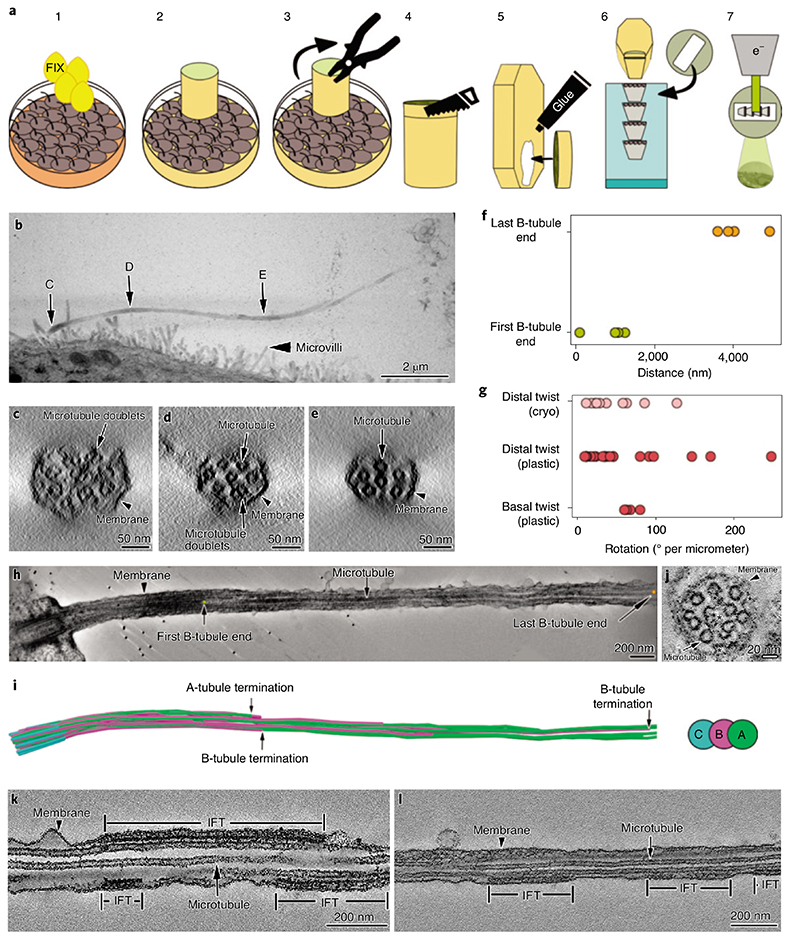
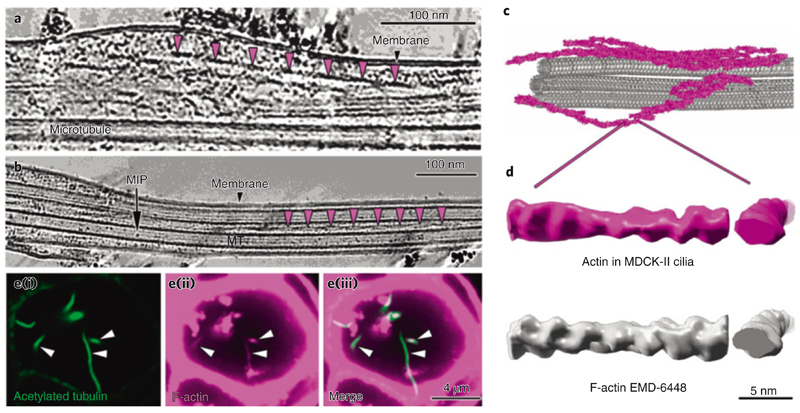
Primary cilia are microtubule-based organelles that are important for signaling and sensing in eukaryotic cells. Unlike the thoroughly studied motile cilia, the three-dimensional architecture and molecular composition of primary cilia are largely unexplored. Yet, studying these aspects is necessary to understand how primary cilia function in health and disease. We developed an enabling method for investigating the structure of primary cilia isolated from MDCK-II cells at molecular resolution by cryo-electron tomography. We show that the textbook '9 + 0' arrangement of microtubule doublets is only present at the primary cilium base. A few microns out, the architecture changes into an unstructured bundle of EB1-decorated microtubules and actin filaments, putting an end to a long debate on the presence or absence of actin filaments in primary cilia. Our work provides a plethora of insights into the molecular structure of primary cilia and offers a methodological framework to study these important organelles.
Figures




Comment in
-
3D architecture of primary cilia by cryo-ET.Nat Rev Nephrol. 2020 Dec;16(12):702. doi: 10.1038/s41581-020-00366-3. Nat Rev Nephrol. 2020. PMID: 33082578 No abstract available.
-
Primary Cilia: A Closer Look at the Antenna of Cells.Curr Biol. 2020 Dec 21;30(24):R1494-R1496. doi: 10.1016/j.cub.2020.10.084. Curr Biol. 2020. PMID: 33352135
References
-
- Badano JL, Mitsuma N, Beales PL, Katsanis N. The ciliopathies: an emerging class of human genetic disorders. Annu Rev Genomics Hum Genet. 2006;7:125–148. - PubMed
-
- Fliegauf M, Benzing T, Omran H. When cilia go bad: cilia defects and ciliopathies. Nat Rev Mol Cell Biol. 2007;8:880–893. - PubMed
-
- Afzelius BA. A human syndrome caused by immotile cilia. Science. 1976;193:317–319. - PubMed
-
- Mitchison HM, Valente EM. Motile and non-motile cilia in human pathology: from function to phenotypes. J Pathol. 2017;241:294–309. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
