

Loss of Extreme Long-Range Enhancers in Human Neural Crest Drives a Craniofacial Disorder
- PMID: 32991838
- PMCID: PMC7655526
- DOI: 10.1016/j.stem.2020.09.001
Loss of Extreme Long-Range Enhancers in Human Neural Crest Drives a Craniofacial Disorder
Abstract
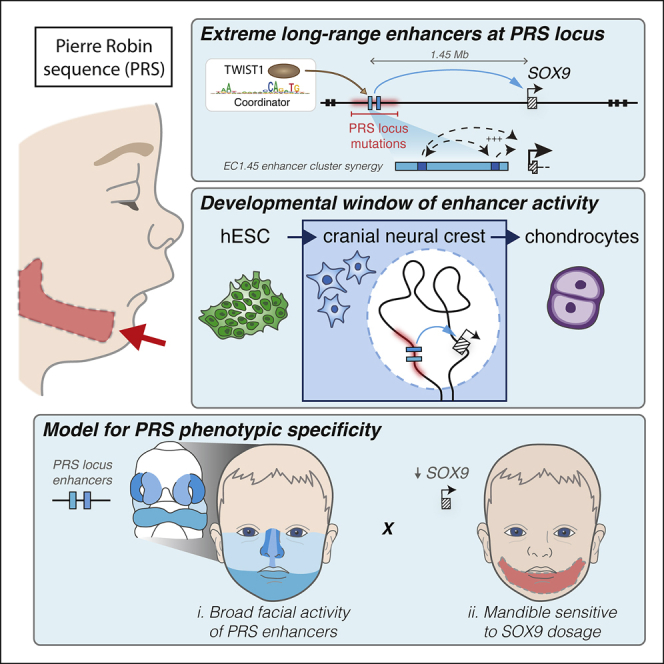
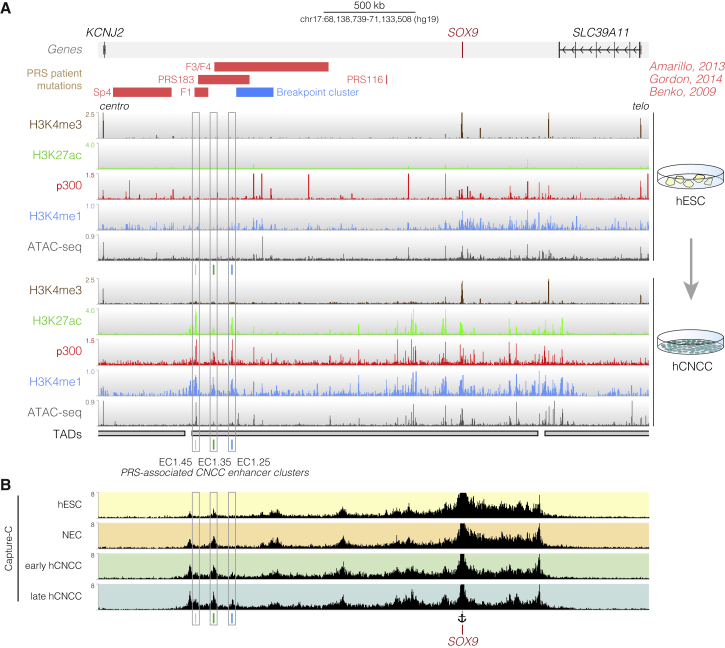
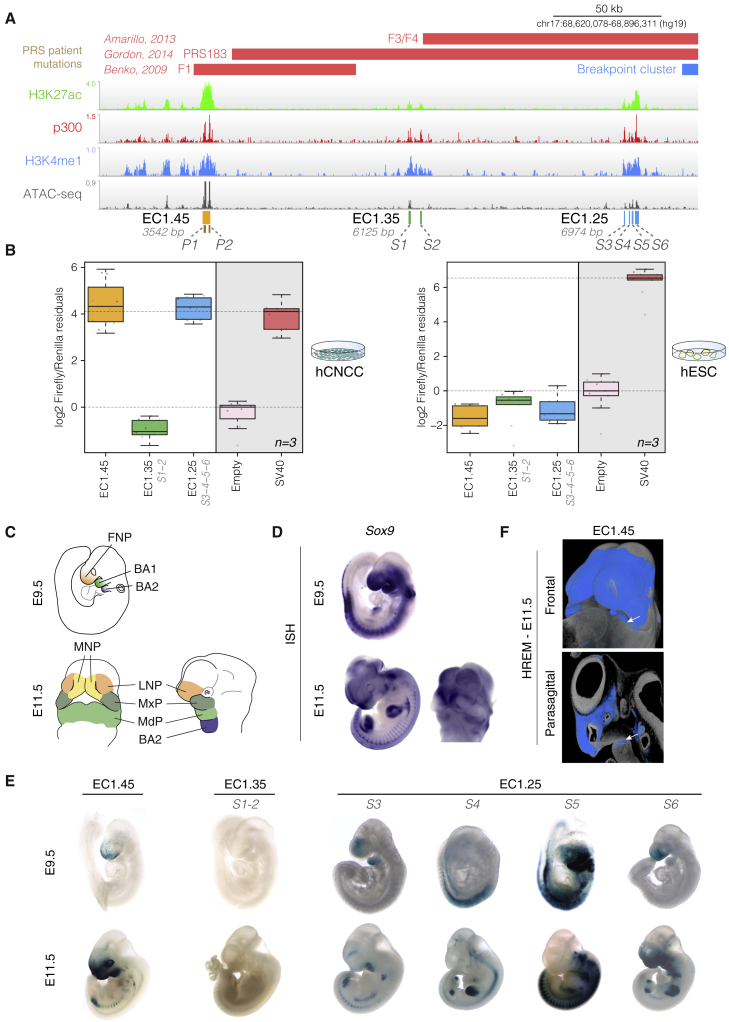
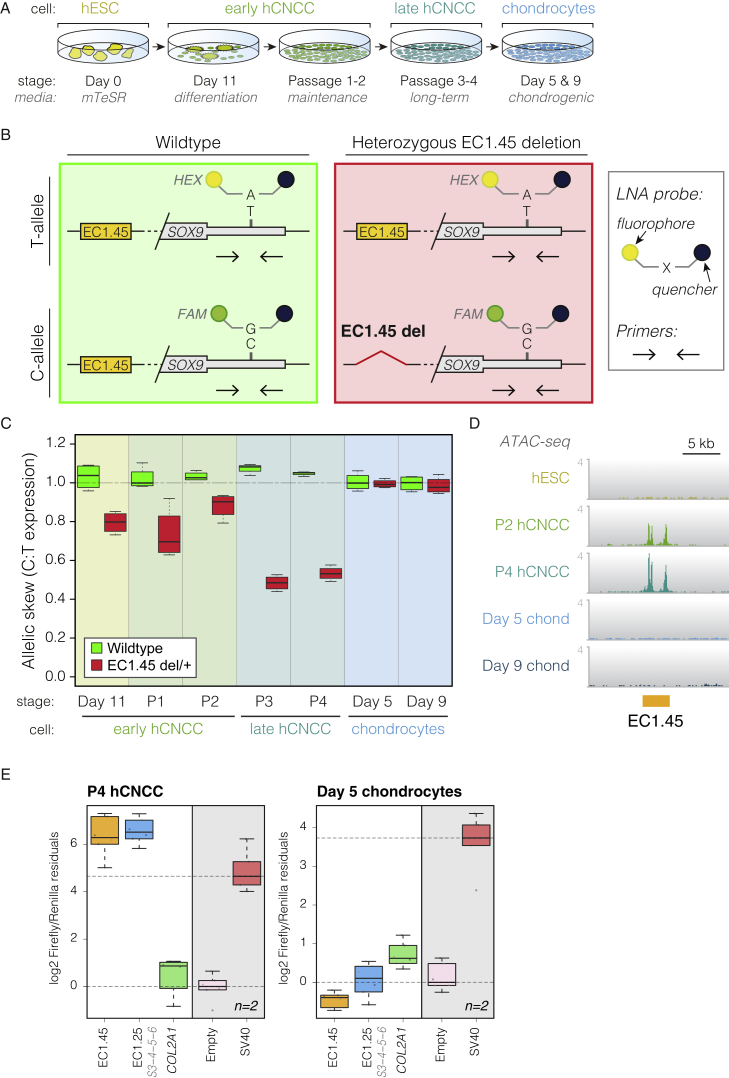
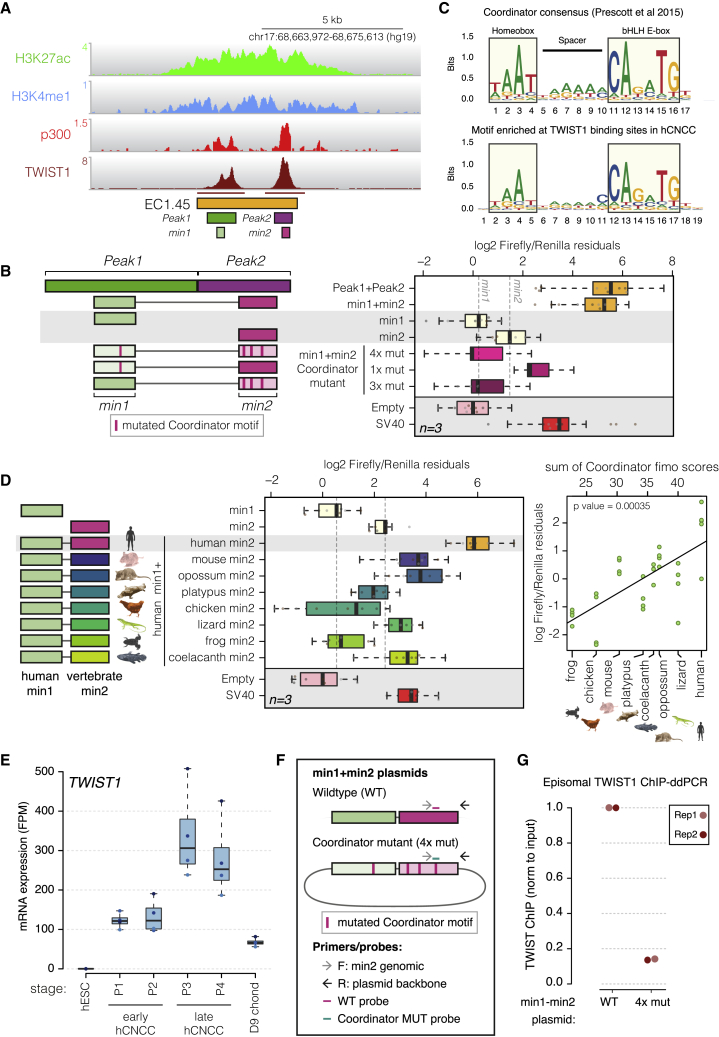
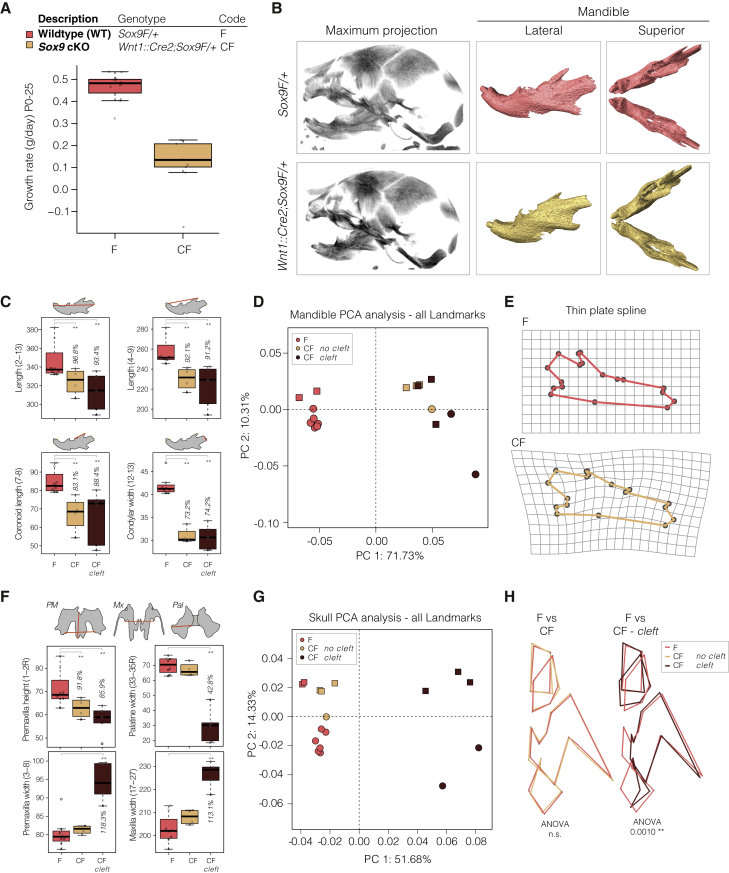
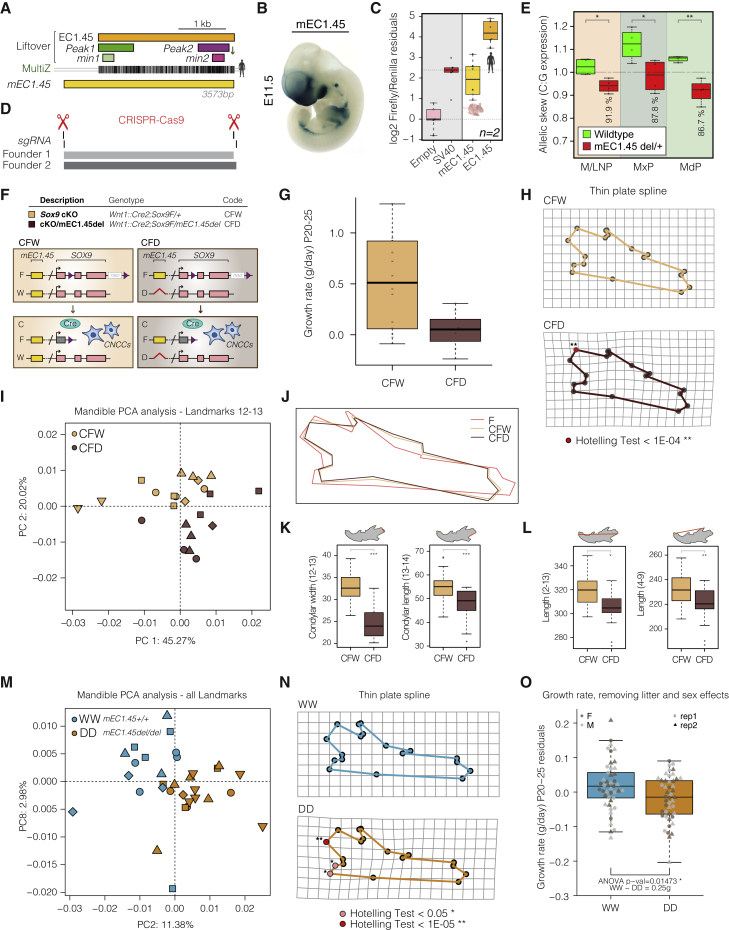
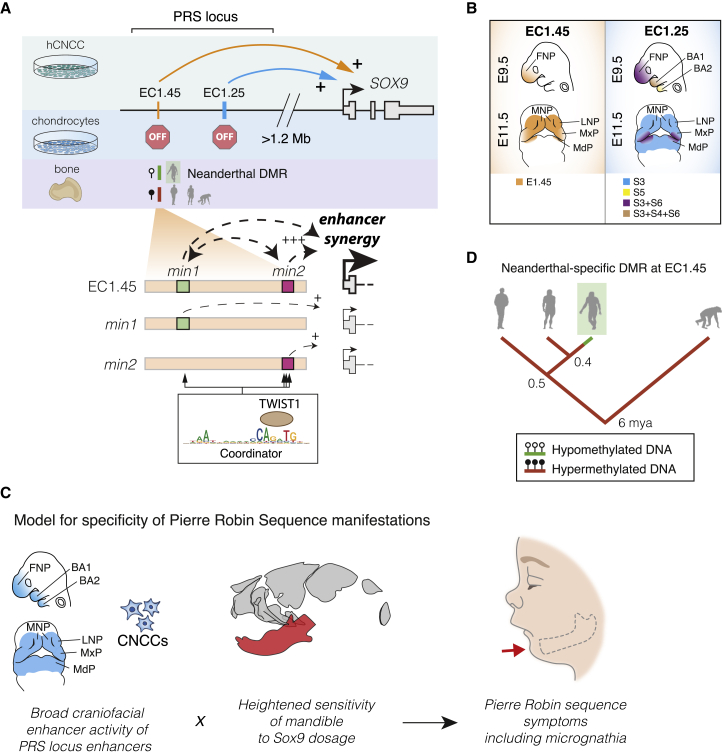
Non-coding mutations at the far end of a large gene desert surrounding the SOX9 gene result in a human craniofacial disorder called Pierre Robin sequence (PRS). Leveraging a human stem cell differentiation model, we identify two clusters of enhancers within the PRS-associated region that regulate SOX9 expression during a restricted window of facial progenitor development at distances up to 1.45 Mb. Enhancers within the 1.45 Mb cluster exhibit highly synergistic activity that is dependent on the Coordinator motif. Using mouse models, we demonstrate that PRS phenotypic specificity arises from the convergence of two mechanisms: confinement of Sox9 dosage perturbation to developing facial structures through context-specific enhancer activity and heightened sensitivity of the lower jaw to Sox9 expression reduction. Overall, we characterize the longest-range human enhancers involved in congenital malformations, directly demonstrate that PRS is an enhanceropathy, and illustrate how small changes in gene expression can lead to morphological variation.
Keywords: Pierre Robin sequence; SOX9; craniofacial; enhancer; enhanceropathy; gene dosage; long-range regulation; neural crest; non-coding mutation; transcription.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests J.W. is a member of the CAMP4 scientific advisory board and ISSCR board of directors. J.R.H. and J.O.J.D. are founders and on the board of directors of Nucleome Theraputics.
Figures
References
-
- Adams D.C., Otárola-Castillo E. Geomorph: An r package for the collection and analysis of geometric morphometric shape data. Methods Ecol. Evol. 2013;4:393–399.
-
- Albertson R.C., Kocher T.D. Genetic and developmental basis of cichlid trophic diversity. Heredity. 2006;97:211–221. - PubMed
-
- Amano O., Doi T., Yamada T., Sasaki A., Sakiyama K., Kanegae H., Kindaichi K. Meckel’s cartilage : Discovery, embryology and evolution -overview of the specificity of meckel’s cartilage- J. Oral Biosci. 2010;52:125–135.
-
- Amarillo I.E., Dipple K.M., Quintero-Rivera F. Familial microdeletion of 17q24.3 upstream of SOX9 is associated with isolated Pierre Robin sequence due to position effect. Am. J. Med. Genet. A. 2013;161A:1167–1172. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DE028599/DE/NIDCR NIH HHS/United States
- U01 DE024430/DE/NIDCR NIH HHS/United States
- R35 GM131757/GM/NIGMS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- U01 DE024427/DE/NIDCR NIH HHS/United States
- T32 GM007276/GM/NIGMS NIH HHS/United States
- MR/R008108/1/MRC_/Medical Research Council/United Kingdom
- F32 DE026950/DE/NIDCR NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- T32 HD007470/HD/NICHD NIH HHS/United States
- MR/N00969X/1/MRC_/Medical Research Council/United Kingdom
- S10 OD018220/OD/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
