

HIV-1 Gag release from yeast reveals ESCRT interaction with the Gag N-terminal protein region
- PMID: 32994219
- PMCID: PMC7939435
- DOI: 10.1074/jbc.RA120.014710
HIV-1 Gag release from yeast reveals ESCRT interaction with the Gag N-terminal protein region
Abstract
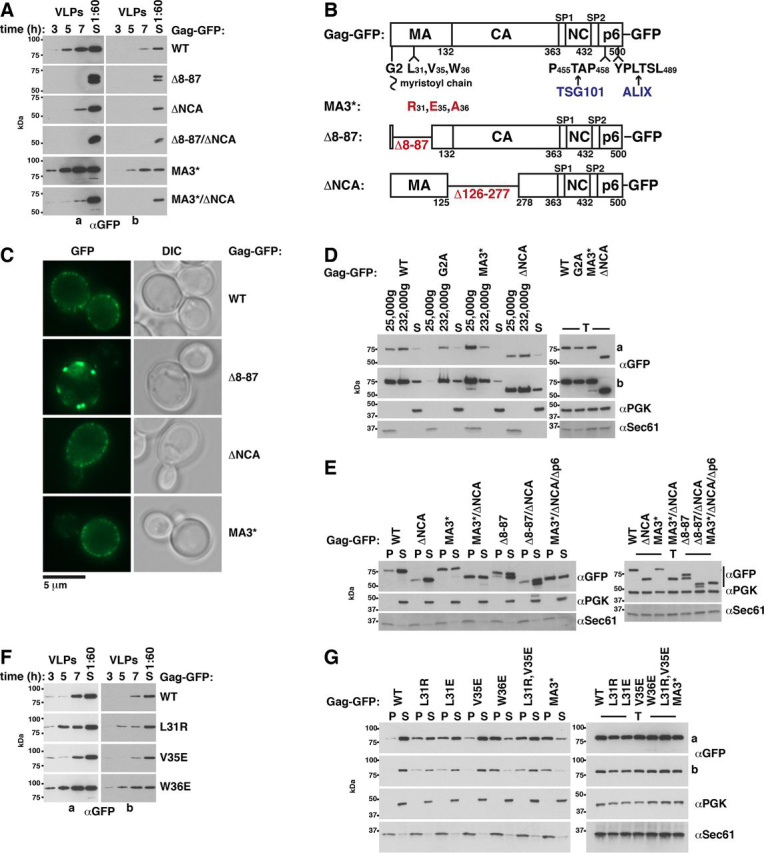
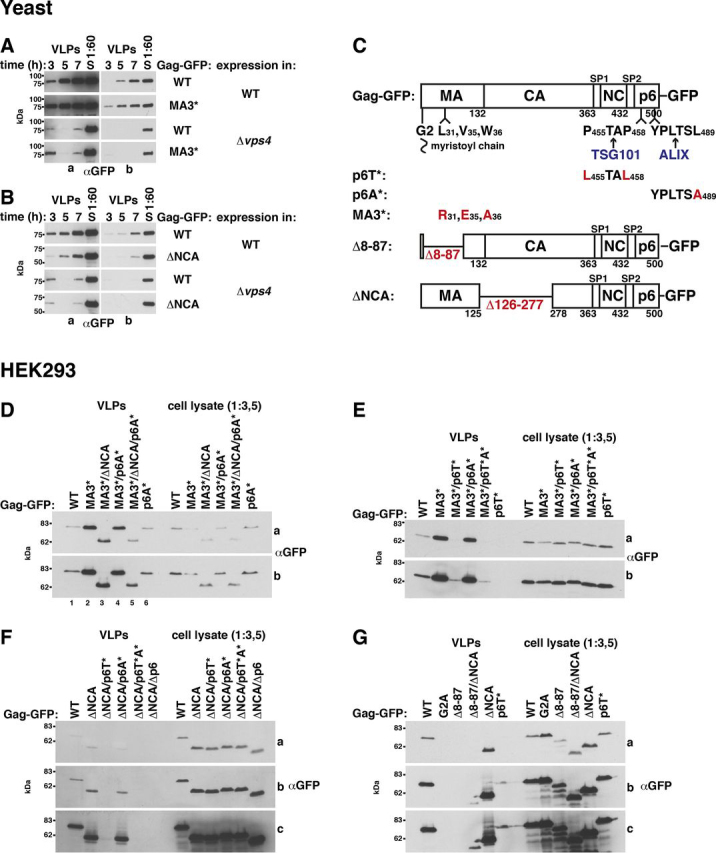
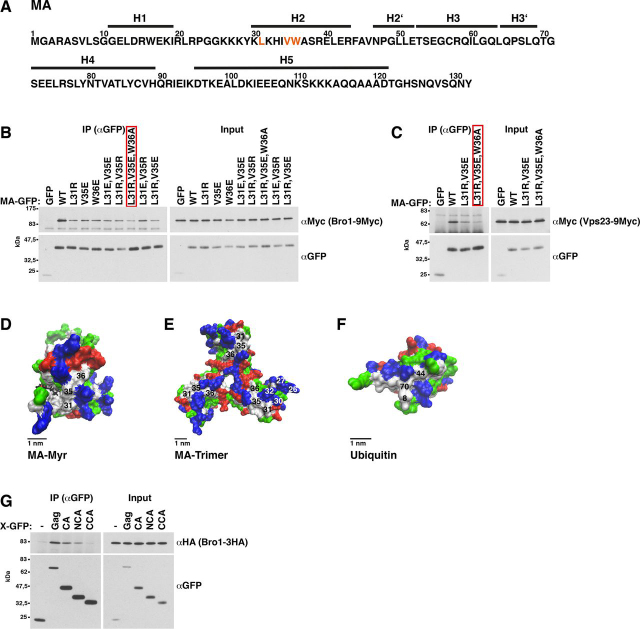
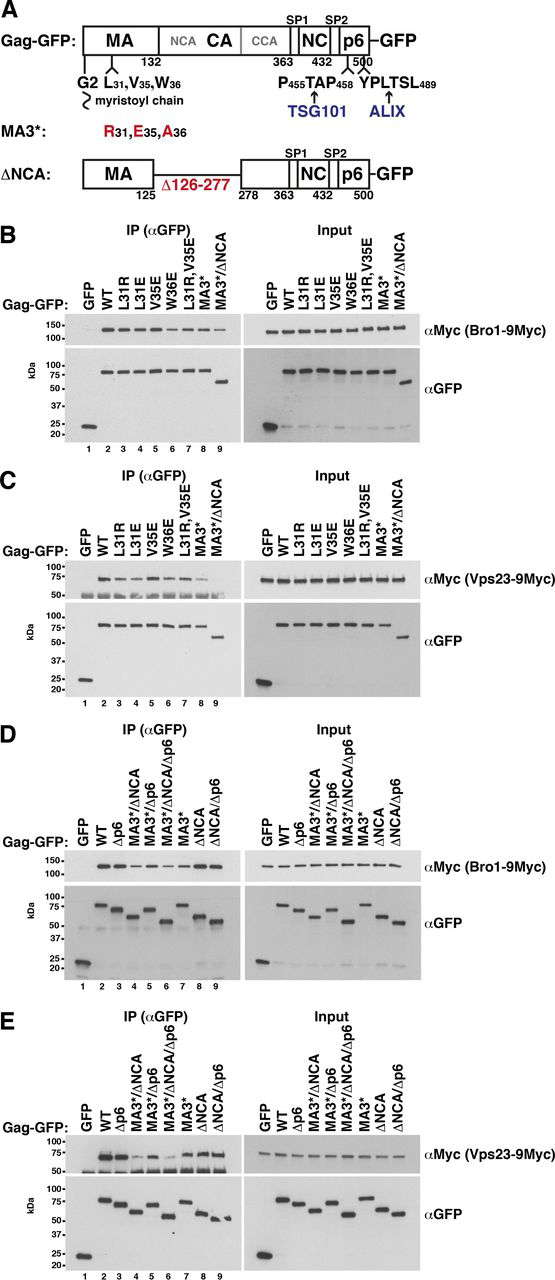
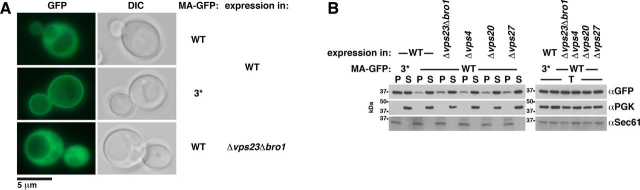
The HIV-1 protein Gag assembles at the plasma membrane and drives virion budding, assisted by the cellular endosomal complex required for transport (ESCRT) proteins. Two ESCRT proteins, TSG101 and ALIX, bind to the Gag C-terminal p6 peptide. TSG101 binding is important for efficient HIV-1 release, but how ESCRTs contribute to the budding process and how their activity is coordinated with Gag assembly is poorly understood. Yeast, allowing genetic manipulation that is not easily available in human cells, has been used to characterize the cellular ESCRT function. Previous work reported Gag budding from yeast spheroplasts, but Gag release was ESCRT-independent. We developed a yeast model for ESCRT-dependent Gag release. We combined yeast genetics and Gag mutational analysis with Gag-ESCRT binding studies and the characterization of Gag-plasma membrane binding and Gag release. With our system, we identified a previously unknown interaction between ESCRT proteins and the Gag N-terminal protein region. Mutations in the Gag-plasma membrane-binding matrix domain that reduced Gag-ESCRT binding increased Gag-plasma membrane binding and Gag release. ESCRT knockout mutants showed that the release enhancement was an ESCRT-dependent effect. Similarly, matrix mutation enhanced Gag release from human HEK293 cells. Release enhancement partly depended on ALIX binding to p6, although binding site mutation did not impair WT Gag release. Accordingly, the relative affinity for matrix compared with p6 in GST-pulldown experiments was higher for ALIX than for TSG101. We suggest that a transient matrix-ESCRT interaction is replaced when Gag binds to the plasma membrane. This step may activate ESCRT proteins and thereby coordinate ESCRT function with virion assembly.
Keywords: ALIX; Bro1; Gag; Saccharomyces cerevisiae; endosomal sorting complexes required for transport (ESCRT); human immunodeficiency virus (HIV); matrix; plasma membrane; virus release; yeast.
© 2020 Meusser et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
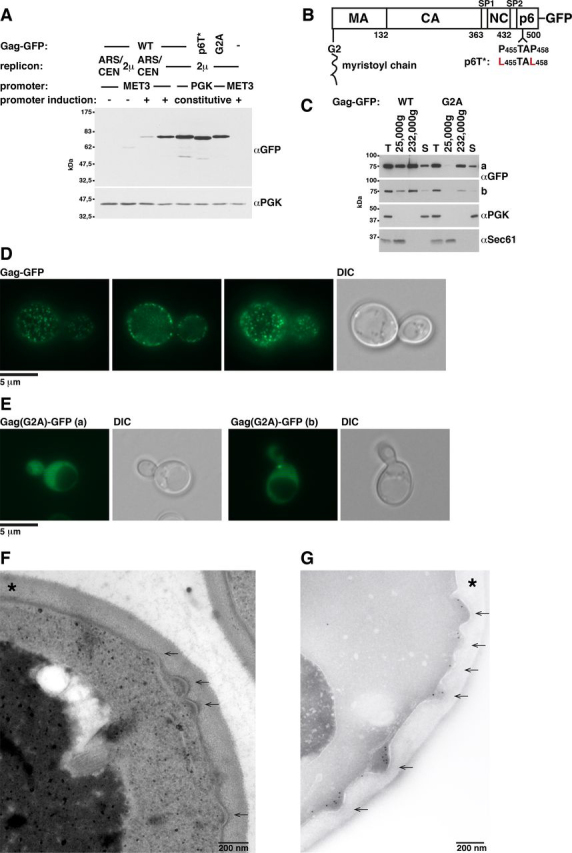
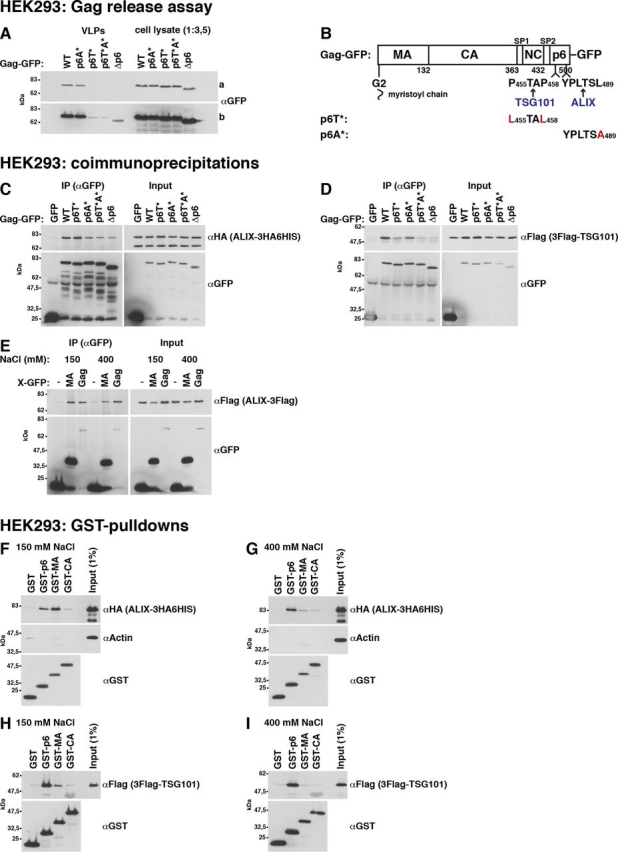
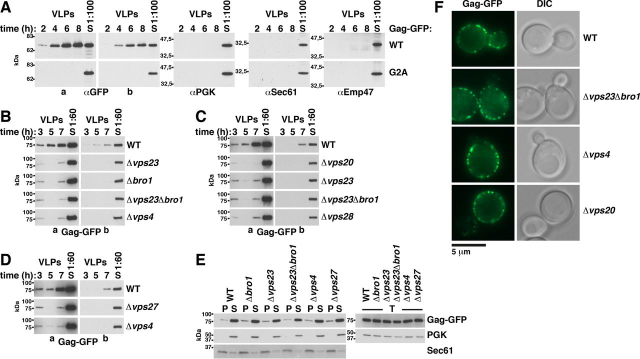
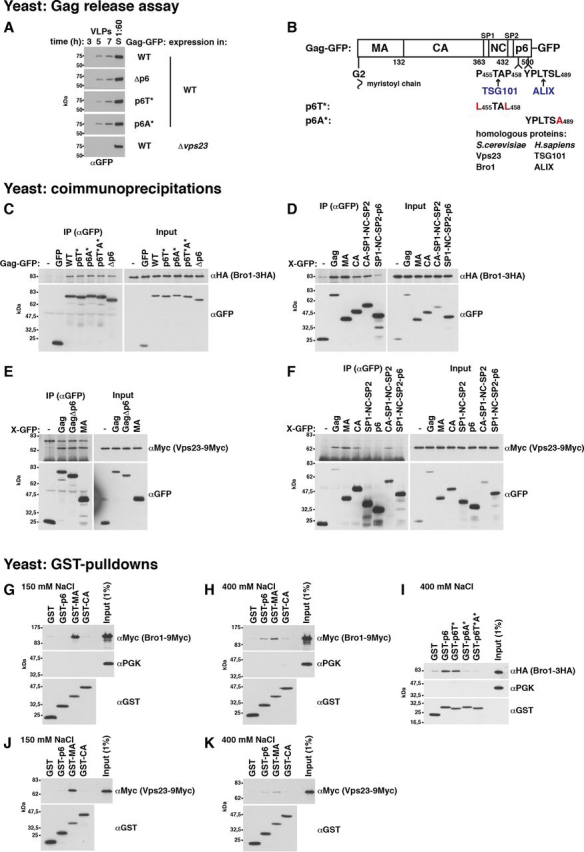
References
Uncited reference
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
