

Kinome Profiling of Primary Endometrial Tumors Using Multiplexed Inhibitor Beads and Mass Spectrometry Identifies SRPK1 as Candidate Therapeutic Target
- PMID: 32994315
- PMCID: PMC7710141
- DOI: 10.1074/mcp.RA120.002012
Kinome Profiling of Primary Endometrial Tumors Using Multiplexed Inhibitor Beads and Mass Spectrometry Identifies SRPK1 as Candidate Therapeutic Target
Abstract
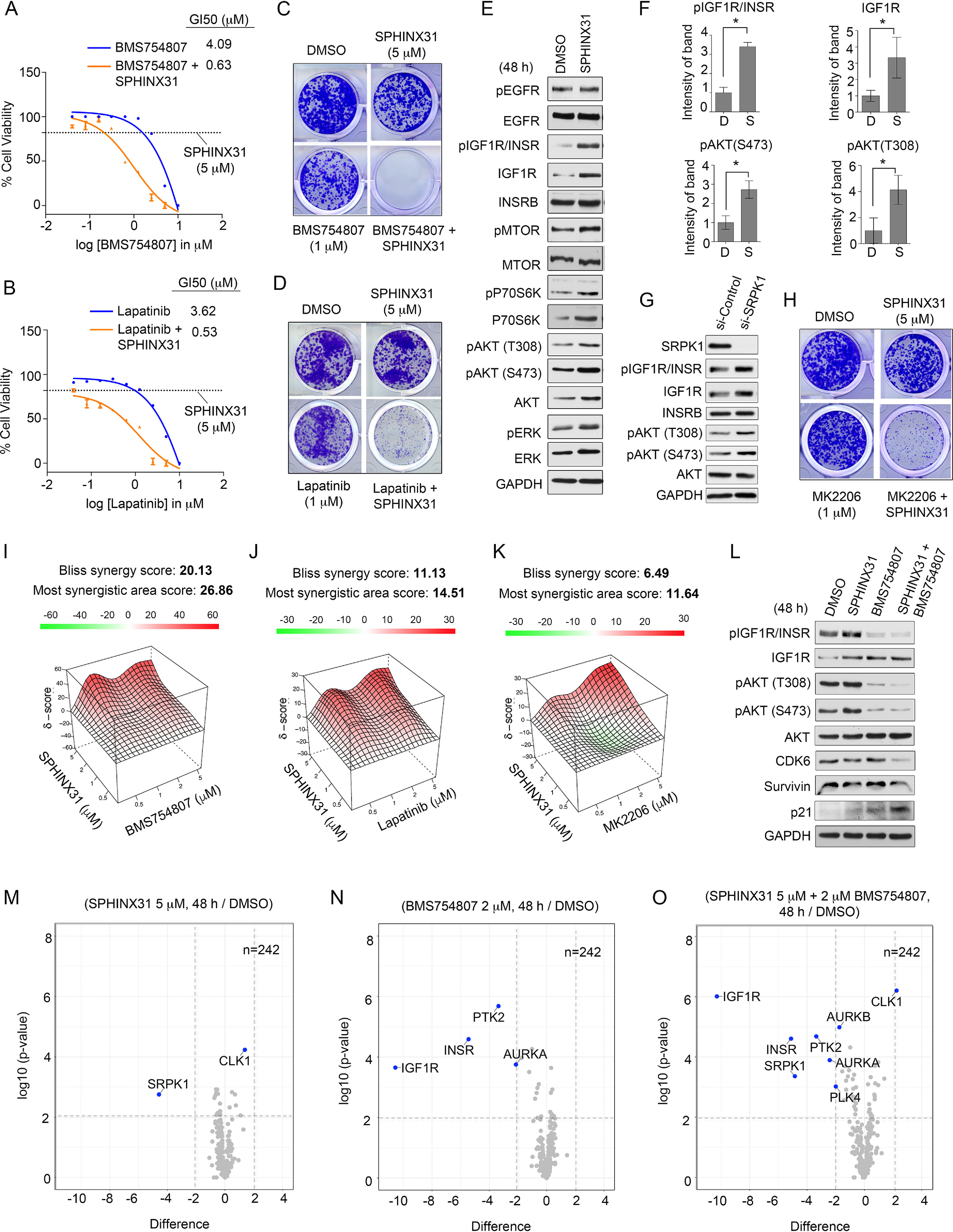
Endometrial carcinoma (EC) is the most common gynecologic malignancy in the United States, with limited effective targeted therapies. Endometrial tumors exhibit frequent alterations in protein kinases, yet only a small fraction of the kinome has been therapeutically explored. To identify kinase therapeutic avenues for EC, we profiled the kinome of endometrial tumors and normal endometrial tissues using Multiplexed Inhibitor Beads and Mass Spectrometry (MIB-MS). Our proteomics analysis identified a network of kinases overexpressed in tumors, including Serine/Arginine-Rich Splicing Factor Kinase 1 (SRPK1). Immunohistochemical (IHC) analysis of endometrial tumors confirmed MIB-MS findings and showed SRPK1 protein levels were highly expressed in endometrioid and uterine serous cancer (USC) histological subtypes. Moreover, querying large-scale genomics studies of EC tumors revealed high expression of SRPK1 correlated with poor survival. Loss-of-function studies targeting SRPK1 in an established USC cell line demonstrated SRPK1 was integral for RNA splicing, as well as cell cycle progression and survival under nutrient deficient conditions. Profiling of USC cells identified a compensatory response to SRPK1 inhibition that involved EGFR and the up-regulation of IGF1R and downstream AKT signaling. Co-targeting SRPK1 and EGFR or IGF1R synergistically enhanced growth inhibition in serous and endometrioid cell lines, representing a promising combination therapy for EC.
Keywords: affinity proteomics; cancer biomarker(s); combination therapies; endometrial carcinoma; kinases; kinome; pathway analysis; splicing; therapeutic targets; tissue proteomics.
© 2020 Kurimchak et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures







References
-
- Siegel R. L., Miller K. D., and Jemal A. (2016) Cancer statistics, 2016. CA Cancer J. Clin. 66, 7–30 - PubMed
-
- Ueda S. M., Kapp D. S., Cheung M. K., Shin J. Y., Osann K., Husain A., Teng N. N., Berek J. S., and Chan J. K. (2008) Trends in demographic and clinical characteristics in women diagnosed with corpus cancer and their potential impact on the increasing number of deaths. Am. J. Obstet. Gynecol. 198, 218, e211–e216 - PubMed
-
- Hecht J. L., and Mutter G. L. (2006) Molecular and pathologic aspects of endometrial carcinogenesis. J. Clin. Oncol. 24, 4783–4791 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
