

Antibiotics and resistance: the two-sided coin of the mycobacterial cell wall
- PMID: 32995684
- PMCID: PMC7502851
- DOI: 10.1016/j.tcsw.2020.100044
Antibiotics and resistance: the two-sided coin of the mycobacterial cell wall
Abstract
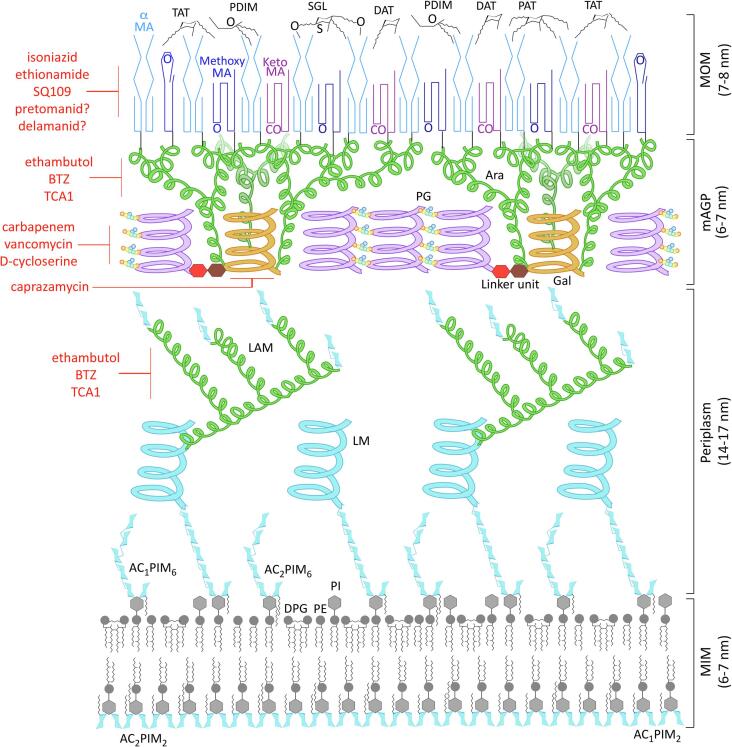
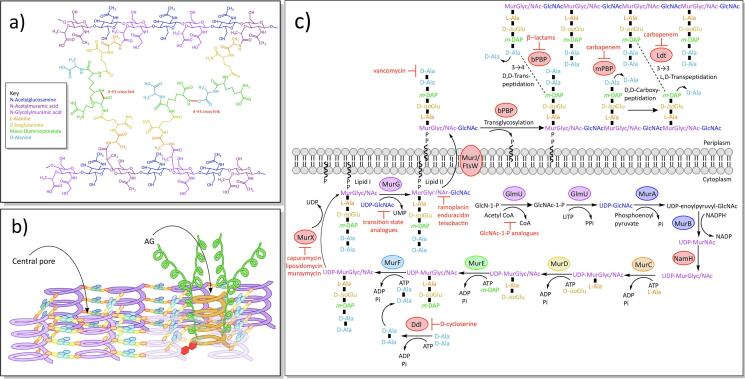
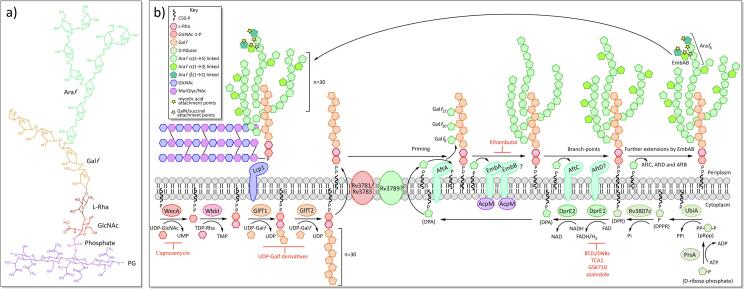
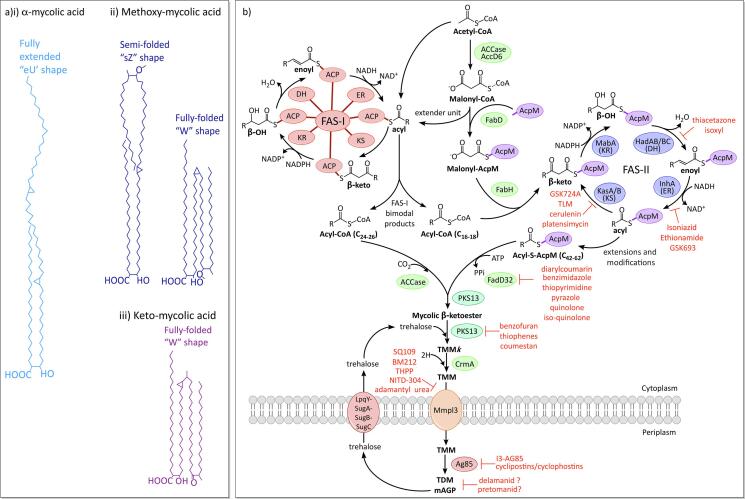
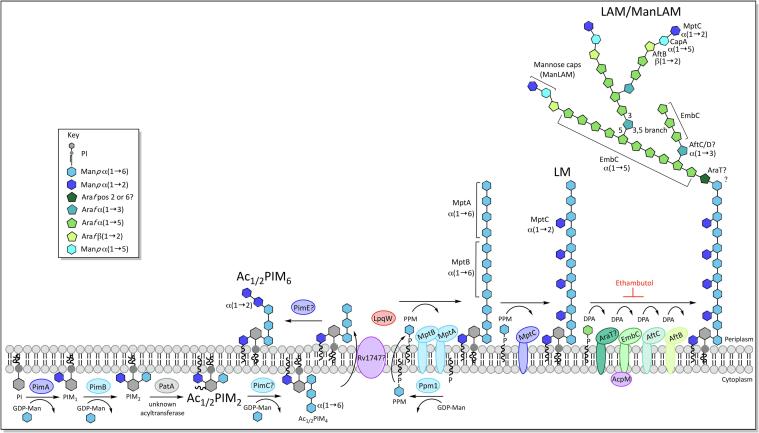
Mycobacterium tuberculosis, the bacterium responsible for tuberculosis, is the global leading cause of mortality from an infectious agent. Part of this success relies on the unique cell wall, which consists of a thick waxy coat with tightly packed layers of complexed sugars, lipids and peptides. This coat provides a protective hydrophobic barrier to antibiotics and the host's defences, while enabling the bacterium to spread efficiently through sputum to infect and survive within the macrophages of new hosts. However, part of this success comes at a cost, with many of the current first- and second-line drugs targeting the enzymes involved in cell wall biosynthesis. The flip side of this coin is that resistance to these drugs develops either in the target enzymes or the activation pathways of the drugs, paving the way for new resistant clinical strains. This review provides a synopsis of the structure and synthesis of the cell wall and the major current drugs and targets, along with any mechanisms of resistance.
Keywords: Arabinogalactan; Cell wall; Lipoarabinomannan; Mycobacterium tuberculosis; Mycolic acids.
© 2020 The Author(s).
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
- Abrahams K.A., Chung C.-W., Ghidelli-Disse S., Rullas J., Rebollo-López M.J., Gurcha S.S., Cox J.A.G., Mendoza A., Jiménez-Navarro E., Martínez-Martínez M.S., Neu M., Shillings A., Homes P., Argyrou A., Casanueva R., Loman N.J., Moynihan P.J., Lelièvre J., Selenski C., Axtman M., Kremer L., Bantscheff M., Angulo-Barturen I., Izquierdo M.C., Cammack N.C., Drewes G., Ballell L., Barros D., Besra G.S., Bates R.H. Identification of KasA as the cellular target of an anti-tubercular scaffold. Nat. Commun. 2016;7:12581. doi: 10.1038/ncomms12581. - DOI - PMC - PubMed
-
- Aggarwal A., Parai M.K., Shetty N., Wallis D., Woolhiser L., Hastings C., Dutta N.K., Galaviz S., Dhakal R.C., Shrestha R., Wakabayashi S., Walpole C., Matthews D., Floyd D., Scullion P., Riley J., Epemolu O., Norval S., Snavely T., Robertson G.T., Rubin E.J., Ioerger T.R., Sirgel F.A., van der Merwe R., van Helden P.D., Keller P., Böttger E.C., Karakousis P.C., Lenaerts A.J., Sacchettini J.C. Development of a novel lead that targets m. tuberculosis polyketide synthase 13. Cell. 2017;170:249–259.e25. doi: 10.1016/j.cell.2017.06.025. - DOI - PMC - PubMed
-
- Alderwick L.J., Radmacher E., Seidel M., Gande R., Hitchen P.G., Morris H.R., Dell A., Sahm H., Eggeling L., Besra G.S. Deletion of Cg-emb in corynebacterianeae leads to a novel truncated cell wall arabinogalactan, whereas inactivation of Cg-ubiA results in an Arabinan-deficient mutant with a cell wall galactan core. J. Biol. Chem. 2005;280:32362–32371. doi: 10.1074/jbc.M506339200. - DOI - PubMed
-
- Alderwick L.J., Dover L.G., Veerapen N., Gurcha S.S., Kremer L., Roper D.L., Pathak A.K., Reynolds R.C., Besra G.S. Expression, purification and characterisation of soluble GlfT and the identification of a novel galactofuranosyltransferase Rv3782 involved in priming GlfT-mediated galactan polymerisation in Mycobacterium tuberculosis. Protein Expr. Purif. 2008;58:332–341. doi: 10.1016/j.pep.2007.11.012. - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
