

This is a preprint.
SARS-CoV-2 nucleocapsid protein forms condensates with viral genomic RNA
- PMID: 32995779
- PMCID: PMC7523105
- DOI: 10.1101/2020.09.14.295824
SARS-CoV-2 nucleocapsid protein forms condensates with viral genomic RNA
Update in
-
SARS-CoV-2 nucleocapsid protein forms condensates with viral genomic RNA.PLoS Biol. 2021 Oct 11;19(10):e3001425. doi: 10.1371/journal.pbio.3001425. eCollection 2021 Oct. PLoS Biol. 2021. PMID: 34634033 Free PMC article.
Abstract
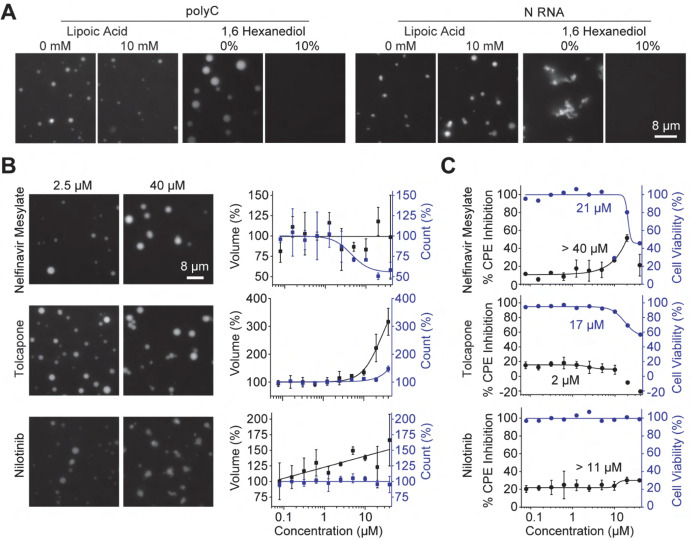
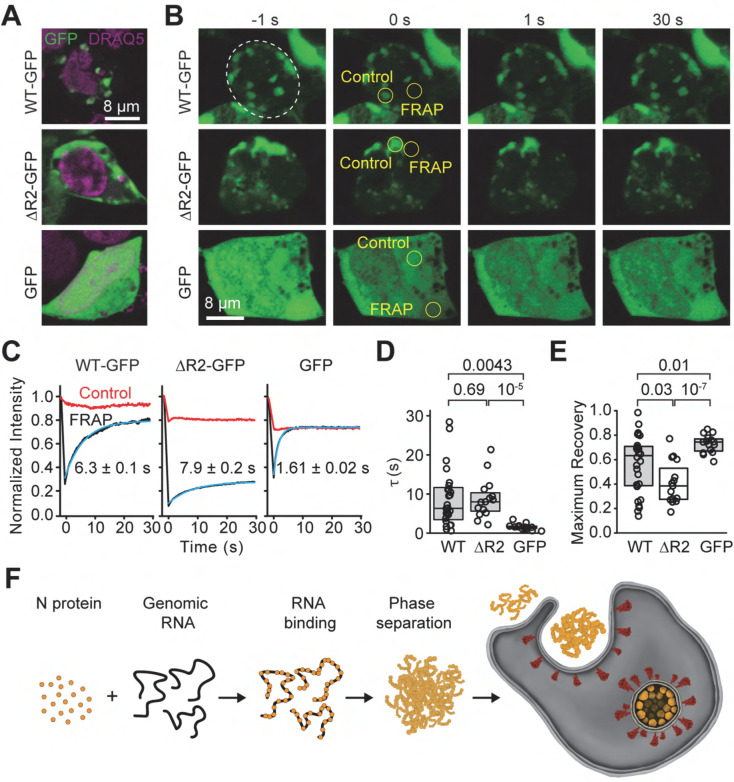
The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection causes COVID-19, a pandemic that seriously threatens global health. SARS-CoV-2 propagates by packaging its RNA genome into membrane enclosures in host cells. The packaging of the viral genome into the nascent virion is mediated by the nucleocapsid (N) protein, but the underlying mechanism remains unclear. Here, we show that the N protein forms biomolecular condensates with viral genomic RNA both in vitro and in mammalian cells. Phase separation is driven, in part, by hydrophobic and electrostatic interactions. While the N protein forms spherical assemblies with unstructured RNA, it forms asymmetric condensates with viral RNA strands that contain secondary structure elements. Cross-linking mass spectrometry identified a region that forms interactions between N proteins in condensates, and truncation of this region disrupts phase separation. We also identified small molecules that alter the formation of N protein condensates. These results suggest that the N protein may utilize biomolecular condensation to package the SARS-CoV-2 RNA genome into a viral particle.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
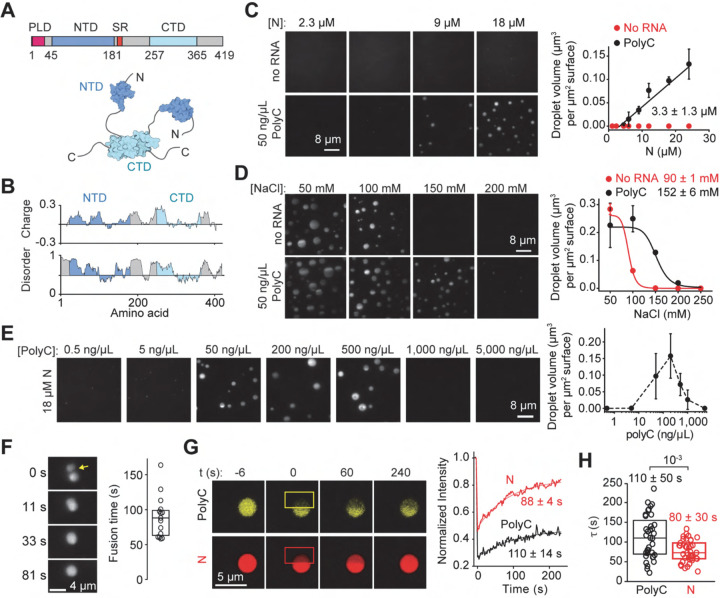
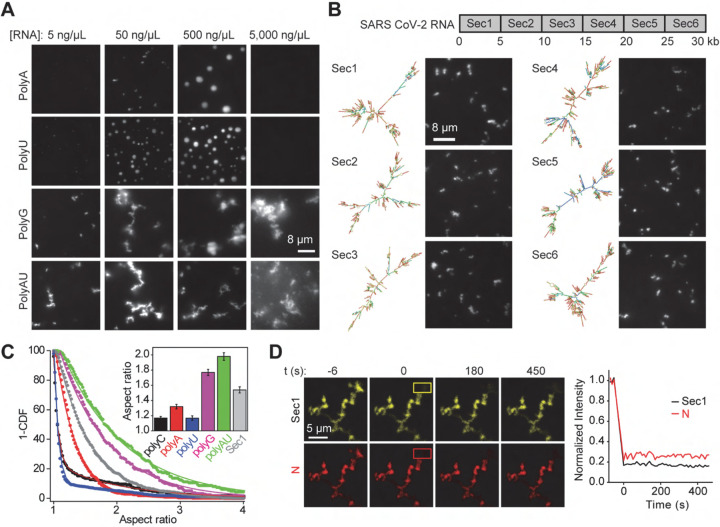
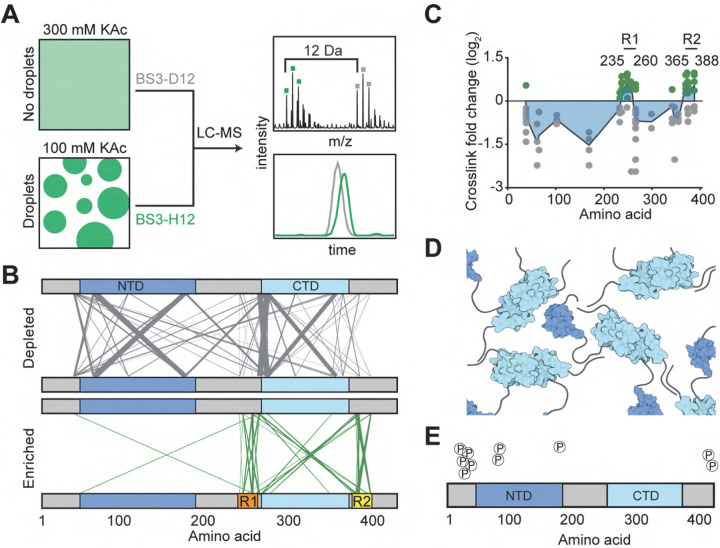
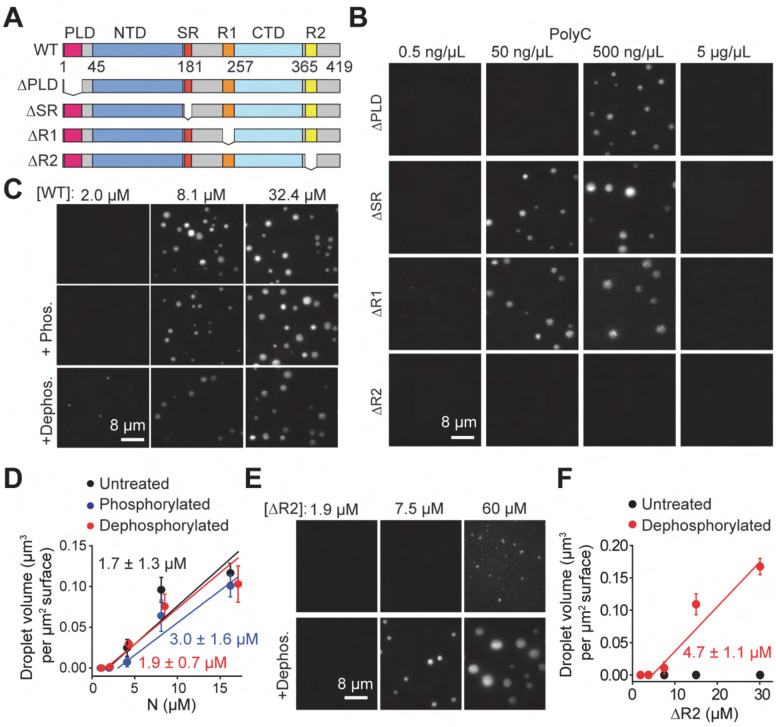
References
-
- Mulligan M. J. et al. , Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 586, 589–593 (2020). - PubMed