

Dux facilitates post-implantation development, but is not essential for zygotic genome activation†
- PMID: 32997106
- PMCID: PMC7786258
- DOI: 10.1093/biolre/ioaa179
Dux facilitates post-implantation development, but is not essential for zygotic genome activation†
Abstract
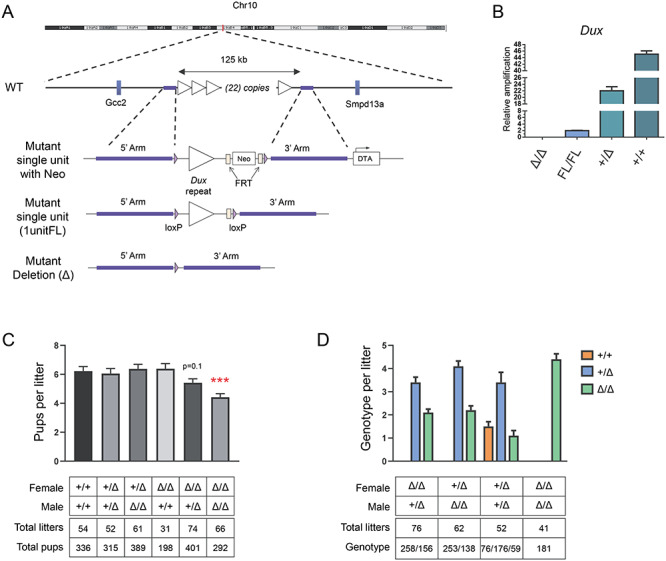
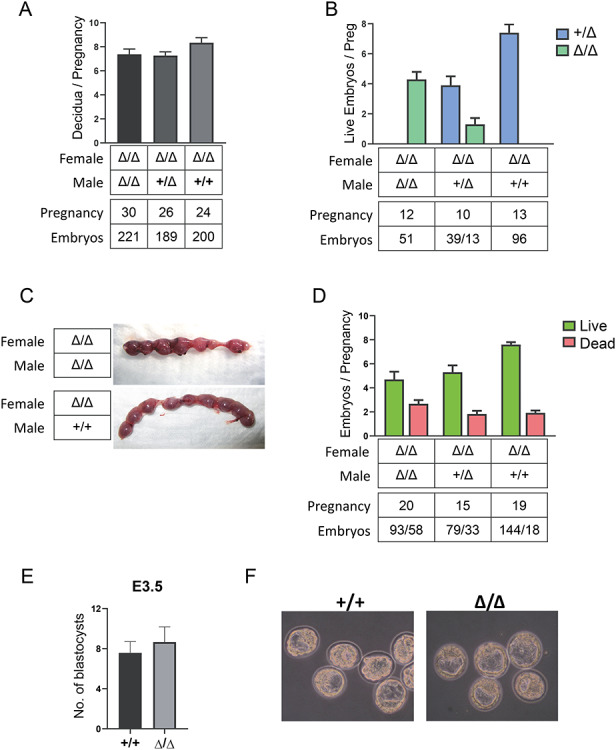
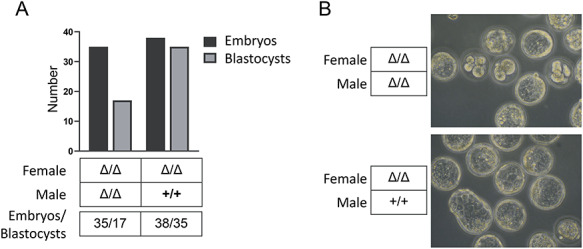
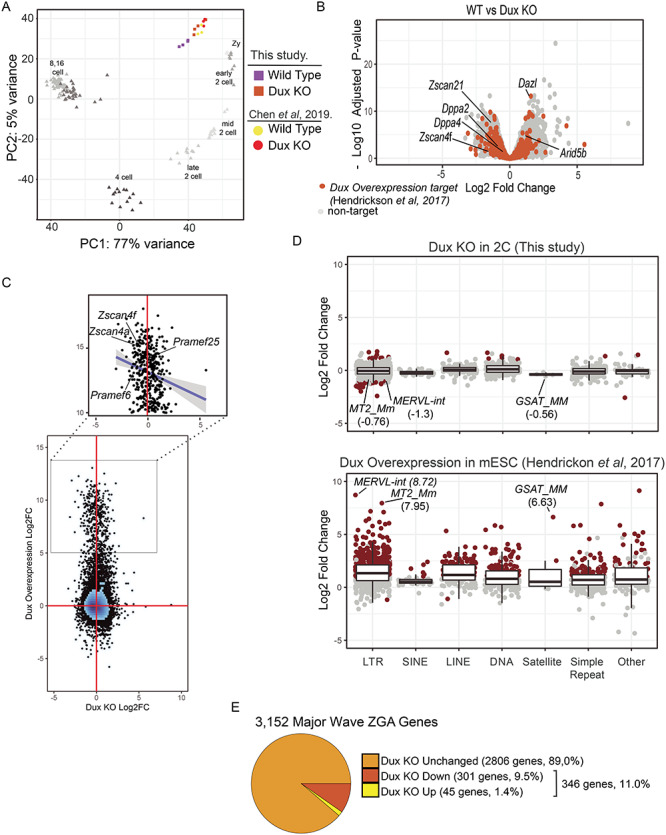
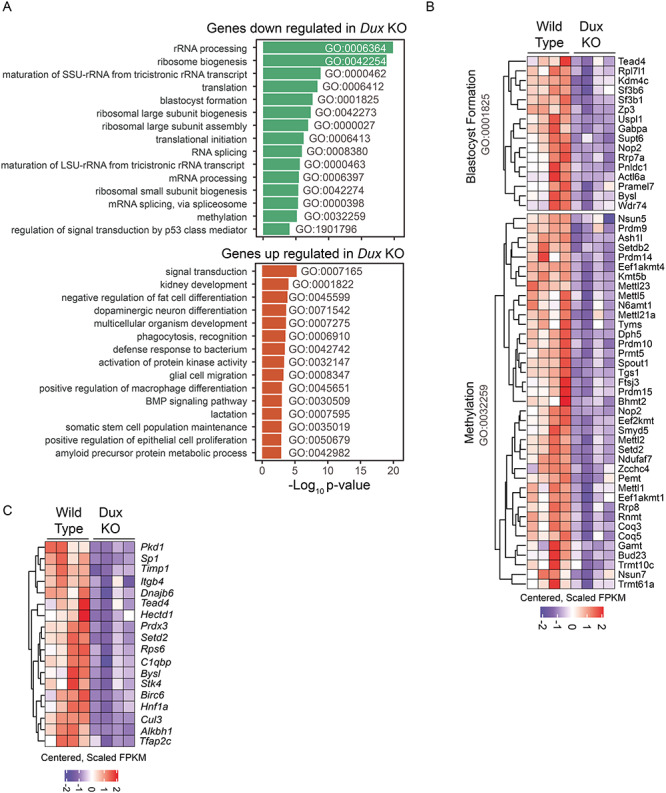
Double homeobox genes are unique to eutherian mammals. It has been proposed that the DUXC clade of the double homeobox gene family, which is present in multicopy long tandem arrays, plays an essential role in zygotic genome activation (ZGA). We generated a deletion of the tandem array encoding the DUXC gene of mouse, Double homeobox (Dux), and found it surprisingly to be homozygous viable and fertile. We characterize the embryonic development and ZGA profile of knockout (KO) embryos, finding that zygotic genome activation still occurs, with only modest alterations in 2-cell embryo gene expression, no defect in in vivo preimplantation development, but an increased likelihood of post-implantation developmental failure, leading to correspondingly smaller litter sizes in the KO strain. While all known 2-cell specific Dux target genes are still expressed in the KO, a subset is expressed at lower levels. These include numerous genes involved in methylation, blastocyst development, and trophectoderm/placental development. We propose that rather than driving ZGA, which is a process common throughout the animal kingdom, DUXC genes facilitate a process unique to eutherian mammals, namely the post-implantation development enabled by an invasive placenta.
Keywords: DUXC; Dux; double homeobox; post-implantation development; zygotic genome activation.
© The Author(s) 2020. Published by Oxford University Press on behalf of Society for the Study of Reproduction. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
