

MYC Promotes Bone Marrow Stem Cell Dysfunction in Fanconi Anemia
- PMID: 32997960
- PMCID: PMC7796920
- DOI: 10.1016/j.stem.2020.09.004
MYC Promotes Bone Marrow Stem Cell Dysfunction in Fanconi Anemia
Abstract
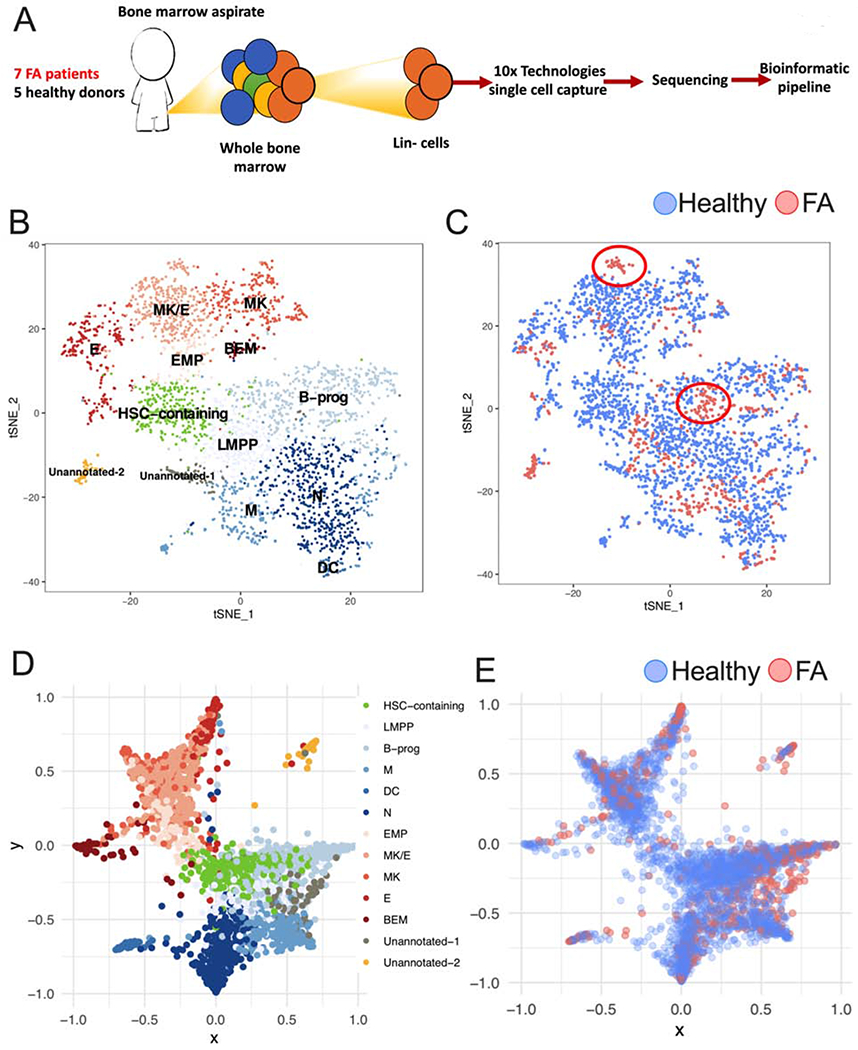
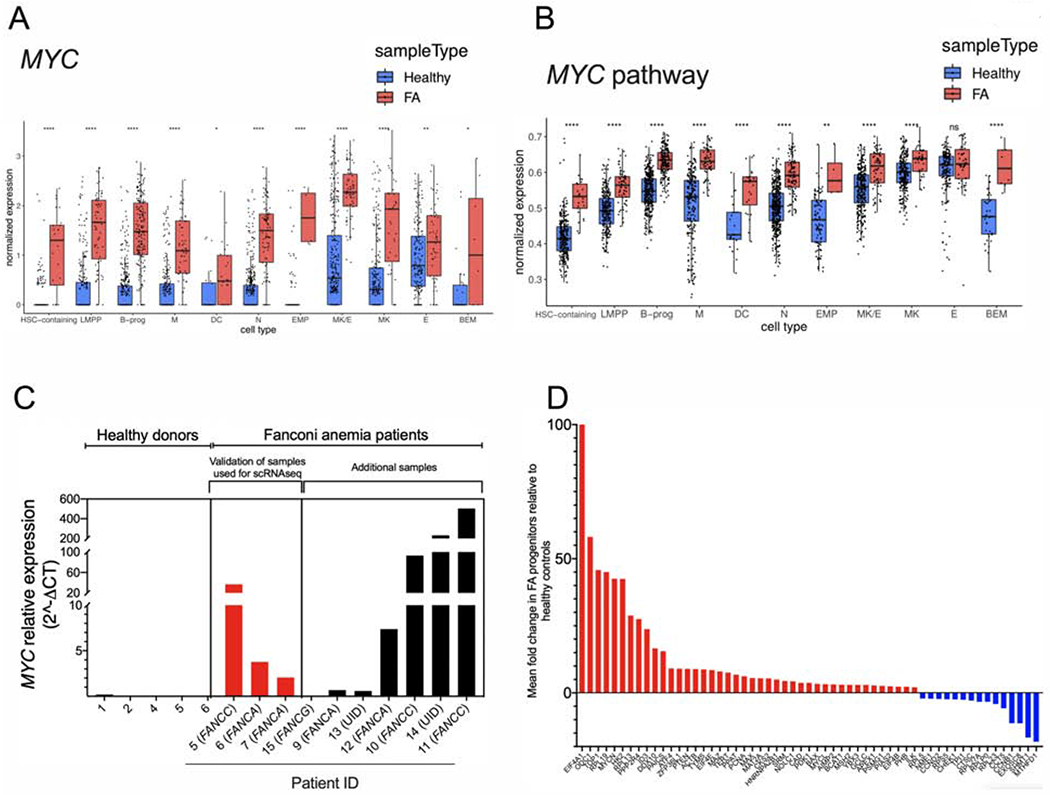
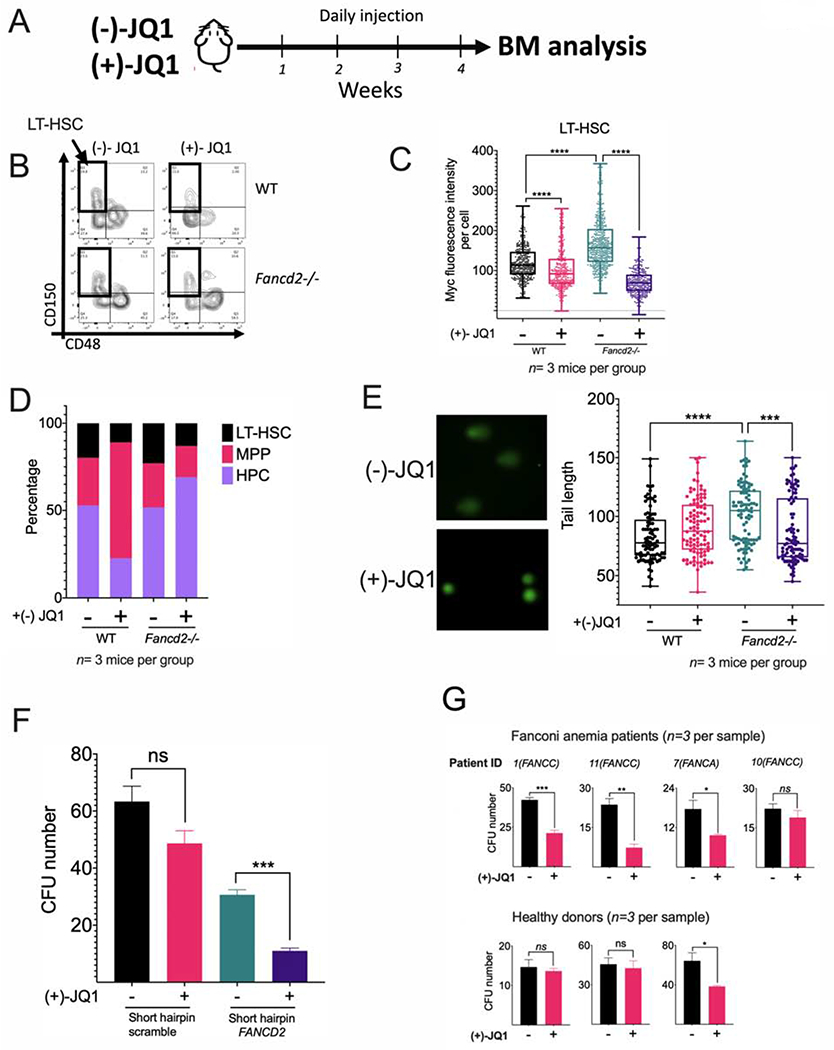
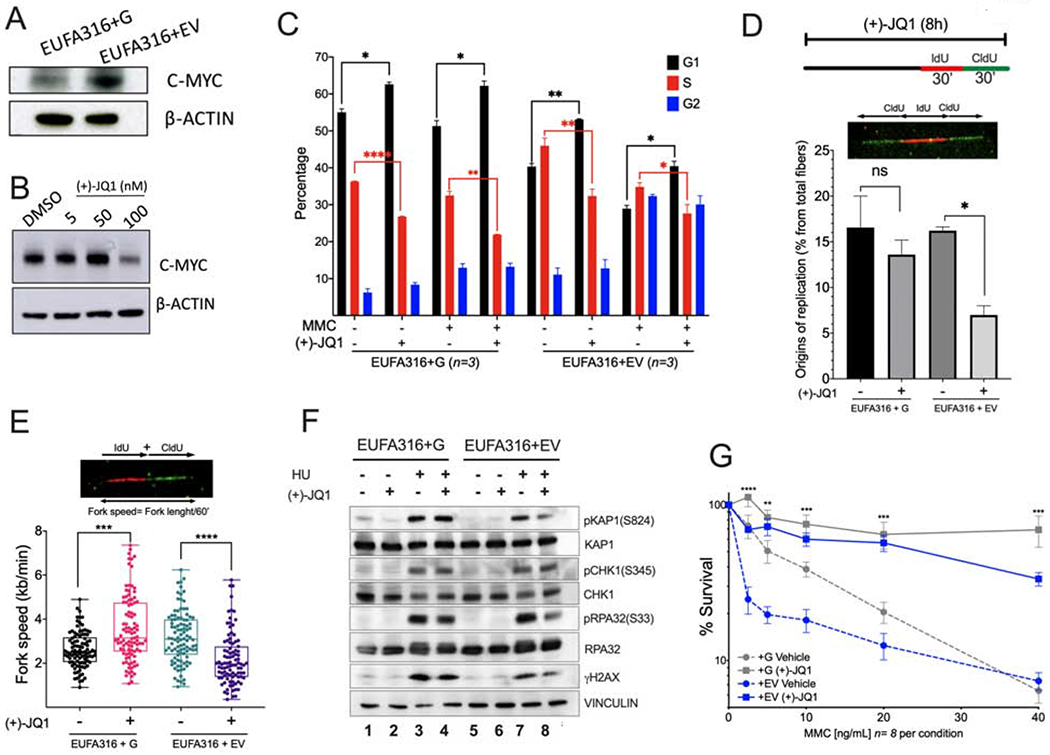
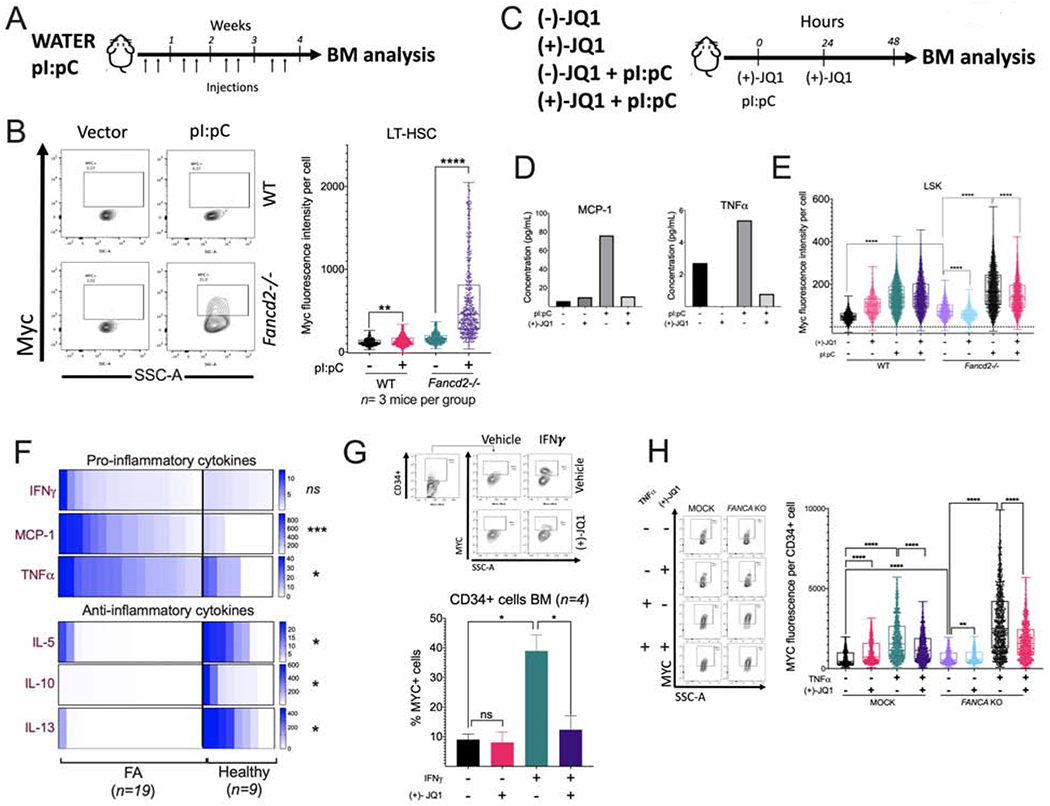
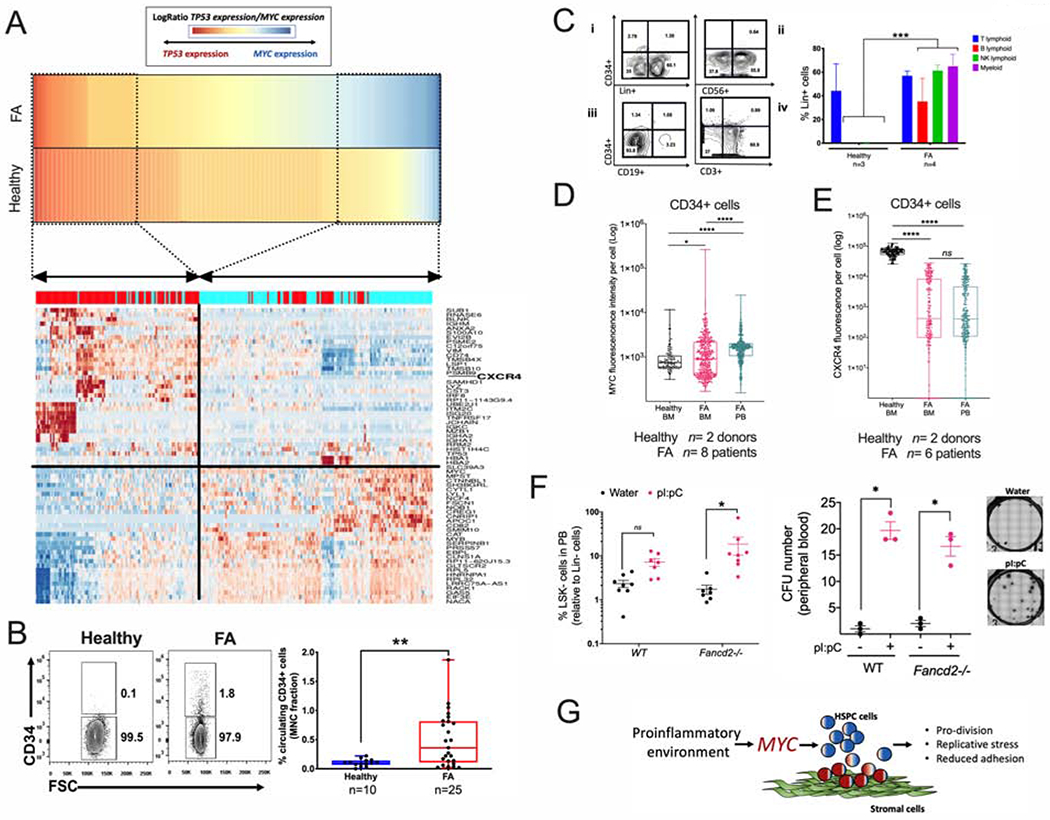
Bone marrow failure (BMF) in Fanconi anemia (FA) patients results from dysfunctional hematopoietic stem and progenitor cells (HSPCs). To identify determinants of BMF, we performed single-cell transcriptome profiling of primary HSPCs from FA patients. In addition to overexpression of p53 and TGF-β pathway genes, we identified high levels of MYC expression. We correspondingly observed coexistence of distinct HSPC subpopulations expressing high levels of TP53 or MYC in FA bone marrow (BM). Inhibiting MYC expression with the BET bromodomain inhibitor (+)-JQ1 reduced the clonogenic potential of FA patient HSPCs but rescued physiological and genotoxic stress in HSPCs from FA mice, showing that MYC promotes proliferation while increasing DNA damage. MYC-high HSPCs showed significant downregulation of cell adhesion genes, consistent with enhanced egress of FA HSPCs from bone marrow to peripheral blood. We speculate that MYC overexpression impairs HSPC function in FA patients and contributes to exhaustion in FA bone marrow.
Keywords: CXCR4; DNA damage; Fanconi anemia; MYC; bone marrow failure; genotoxic stress; hematopoietic stem cells; p53; physiological stress; single-cell RNA sequencing.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures

Comment in
-
Tipping the Scale: MYC Gains Weight in Fanconi Anemia Bone Marrow Failure Progression.Cell Stem Cell. 2021 Jan 7;28(1):8-9. doi: 10.1016/j.stem.2020.12.013. Cell Stem Cell. 2021. PMID: 33417873
References
-
- ADAMO A, COLLIS SJ, ADELMAN CA, SILVA N, HOREJSI Z, WARD JD, MARTINEZ-PEREZ E, BOULTON SJ & LA VOLPE A 2010. Preventing nonhomologous end joining suppresses DNA repair defects of Fanconi anemia. Mol Cell, 39, 25–35. - PubMed
-
- BAHR C, VON PALESKE L, USLU VV, REMESEIRO S, TAKAYAMA N, NG SW, MURISON A, LANGENFELD K, PETRETICH M, SCOGNAMIGLIO R, ZEISBERGER P, BENK AS, AMIT I, ZANDSTRA PW, LUPIEN M, DICK JE, TRUMPP A & SPITZ F 2018. A Myc enhancer cluster regulates normal and leukaemic haematopoietic stem cell hierarchies. Nature, 553, 515–520. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
