

Innovations present in the primate interneuron repertoire
- PMID: 32999462
- PMCID: PMC7957574
- DOI: 10.1038/s41586-020-2781-z
Innovations present in the primate interneuron repertoire
Erratum in
-
Author Correction: Innovations present in the primate interneuron repertoire.Nature. 2020 Dec;588(7837):E17. doi: 10.1038/s41586-020-2874-8. Nature. 2020. PMID: 33230336
Abstract
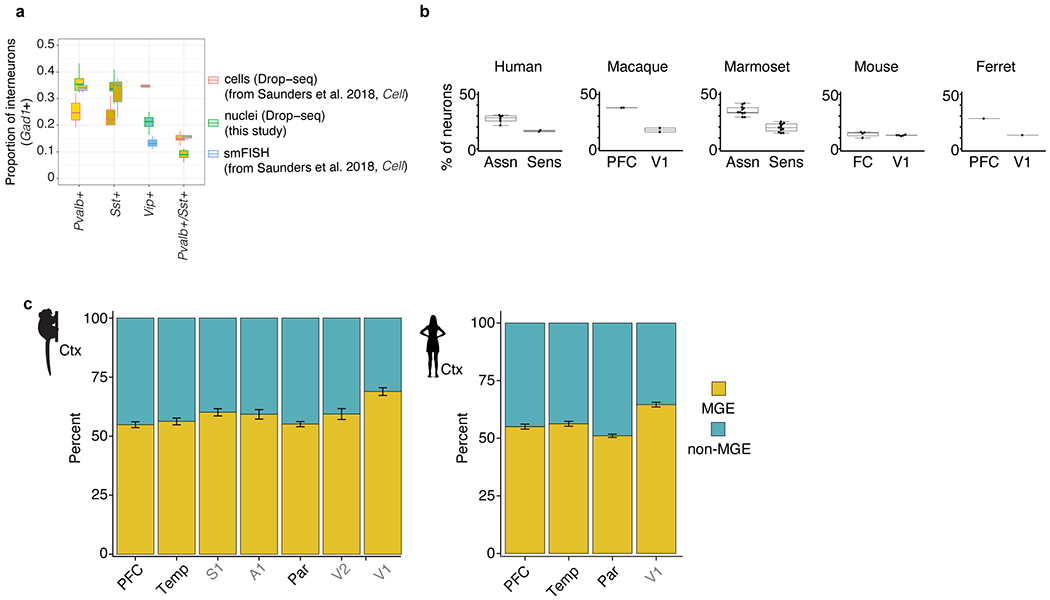
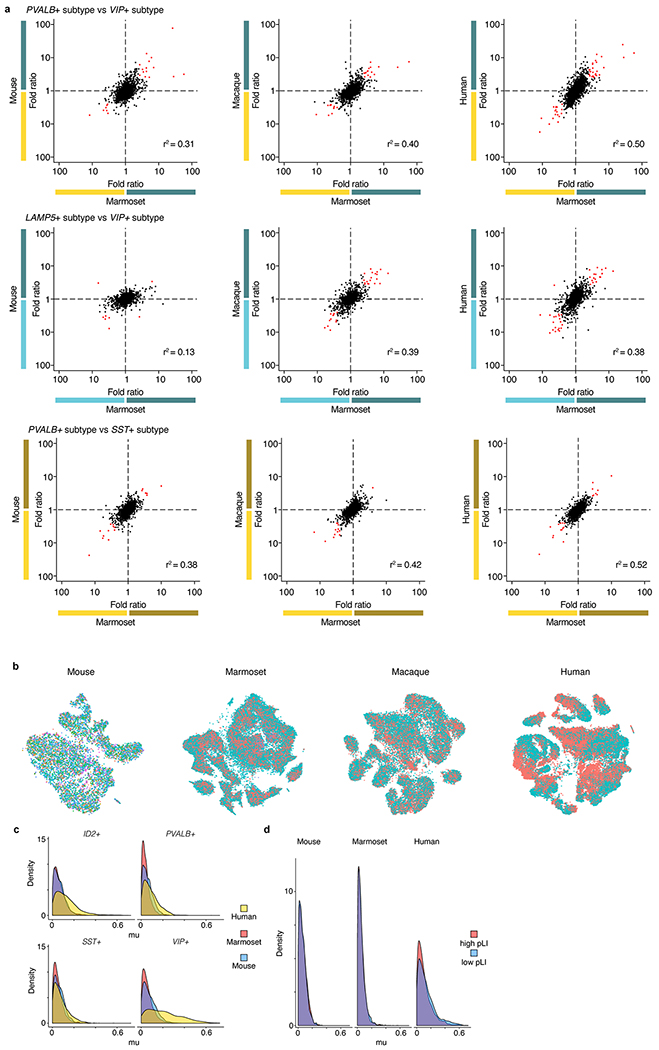
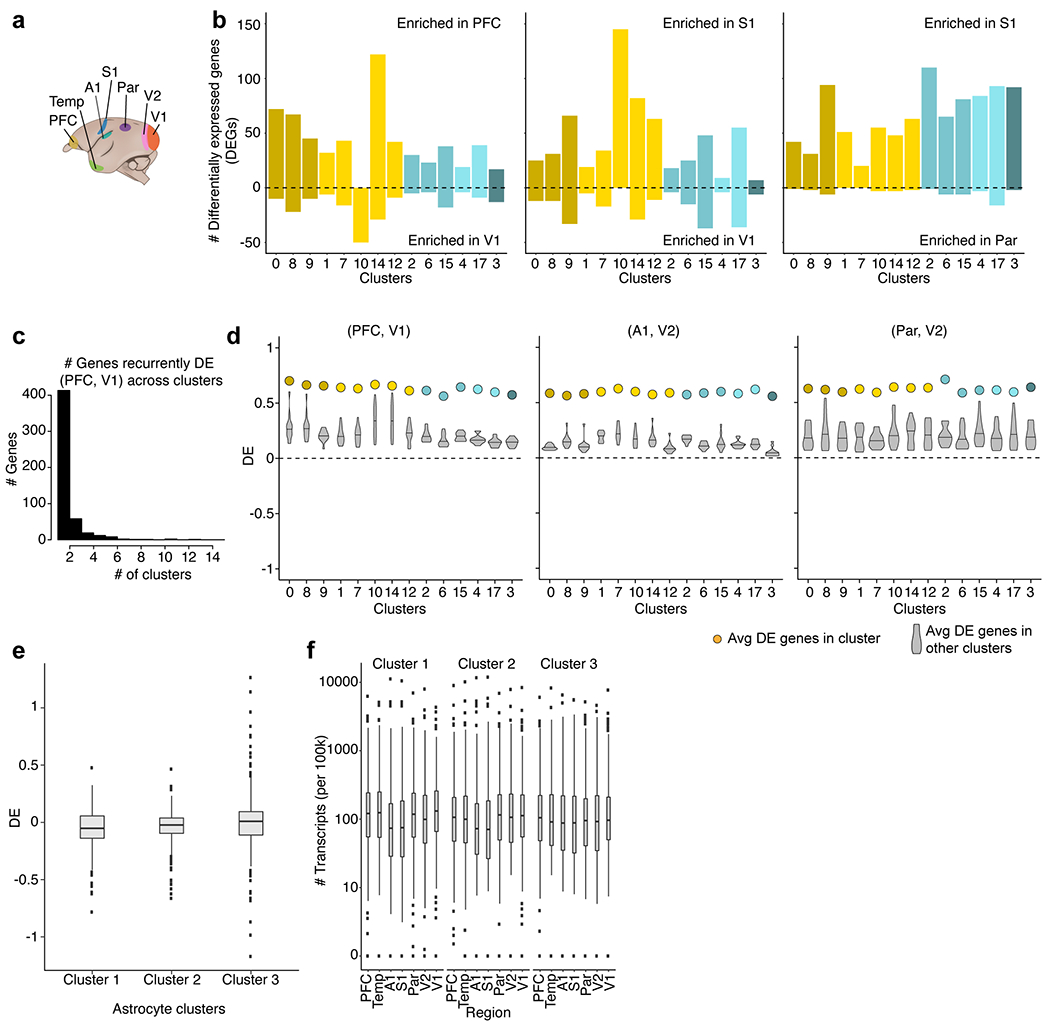
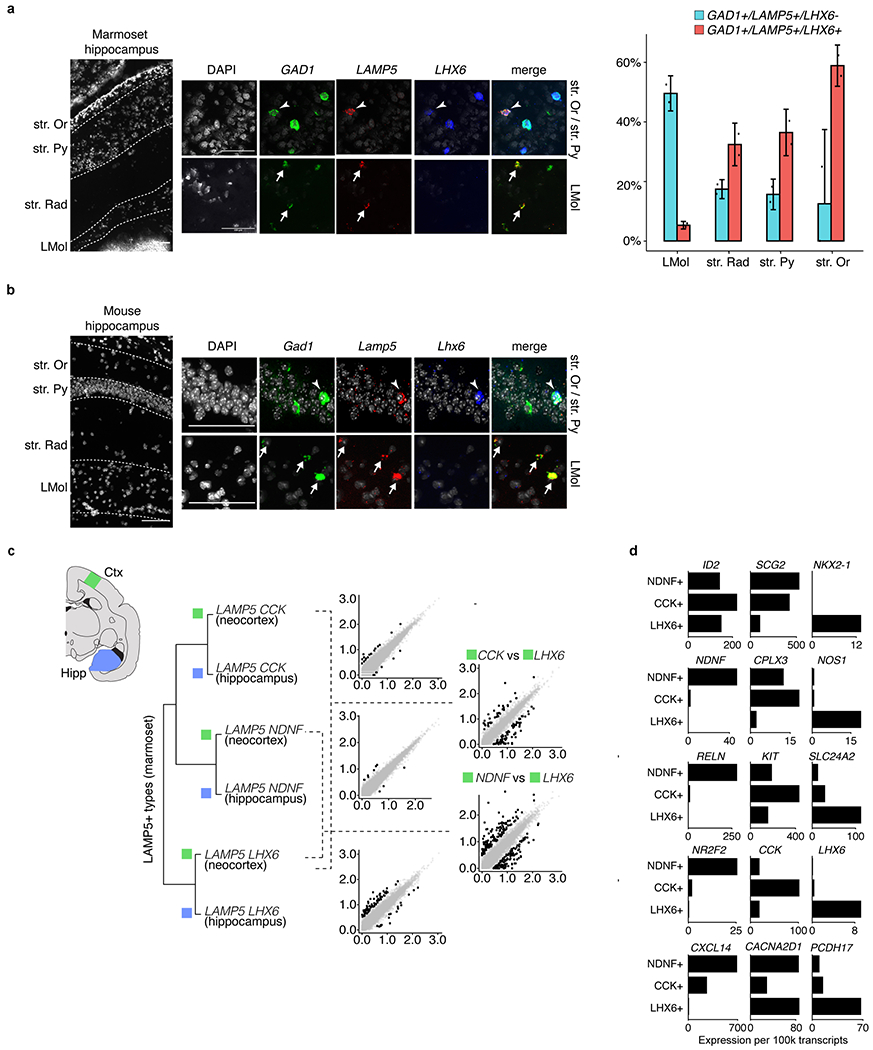
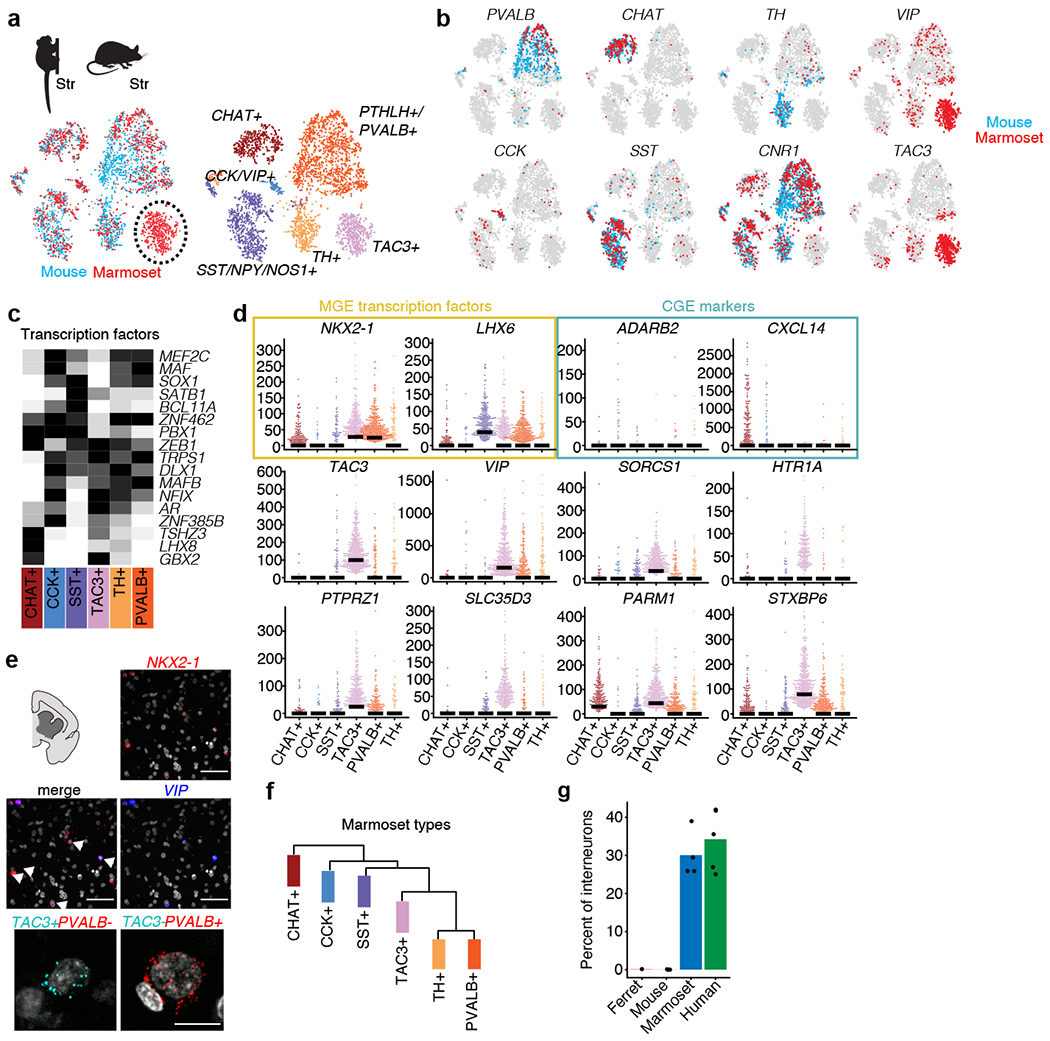
Primates and rodents, which descended from a common ancestor around 90 million years ago1, exhibit profound differences in behaviour and cognitive capacity; the cellular basis for these differences is unknown. Here we use single-nucleus RNA sequencing to profile RNA expression in 188,776 individual interneurons across homologous brain regions from three primates (human, macaque and marmoset), a rodent (mouse) and a weasel (ferret). Homologous interneuron types-which were readily identified by their RNA-expression patterns-varied in abundance and RNA expression among ferrets, mice and primates, but varied less among primates. Only a modest fraction of the genes identified as 'markers' of specific interneuron subtypes in any one species had this property in another species. In the primate neocortex, dozens of genes showed spatial expression gradients among interneurons of the same type, which suggests that regional variation in cortical contexts shapes the RNA expression patterns of adult neocortical interneurons. We found that an interneuron type that was previously associated with the mouse hippocampus-the 'ivy cell', which has neurogliaform characteristics-has become abundant across the neocortex of humans, macaques and marmosets but not mice or ferrets. We also found a notable subcortical innovation: an abundant striatal interneuron type in primates that had no molecularly homologous counterpart in mice or ferrets. These interneurons expressed a unique combination of genes that encode transcription factors, receptors and neuropeptides and constituted around 30% of striatal interneurons in marmosets and humans.
Conflict of interest statement
Figures









Similar articles
-
Peripuberty Is a Sensitive Period for Prefrontal Parvalbumin Interneuron Activity to Impact Adult Cognitive Flexibility.Dev Neurosci. 2025;47(2):127-138. doi: 10.1159/000539584. Epub 2024 Jun 3. Dev Neurosci. 2025. PMID: 38830346 Free PMC article.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
Signs and symptoms to determine if a patient presenting in primary care or hospital outpatient settings has COVID-19.Cochrane Database Syst Rev. 2022 May 20;5(5):CD013665. doi: 10.1002/14651858.CD013665.pub3. Cochrane Database Syst Rev. 2022. PMID: 35593186 Free PMC article.
-
Interventions for prevention of herpes simplex labialis (cold sores on the lips).Cochrane Database Syst Rev. 2015 Aug 7;2015(8):CD010095. doi: 10.1002/14651858.CD010095.pub2. Cochrane Database Syst Rev. 2015. PMID: 26252373 Free PMC article.
-
Sertindole for schizophrenia.Cochrane Database Syst Rev. 2005 Jul 20;2005(3):CD001715. doi: 10.1002/14651858.CD001715.pub2. Cochrane Database Syst Rev. 2005. PMID: 16034864 Free PMC article.
Cited by
-
Cellular-resolution gene expression profiling in the neonatal marmoset brain reveals dynamic species- and region-specific differences.Proc Natl Acad Sci U S A. 2021 May 4;118(18):e2020125118. doi: 10.1073/pnas.2020125118. Proc Natl Acad Sci U S A. 2021. PMID: 33903237 Free PMC article.
-
A scalable, high-throughput neural development platform identifies shared impact of ASD genes on cell fate and differentiation.bioRxiv [Preprint]. 2024 Sep 26:2024.09.25.614184. doi: 10.1101/2024.09.25.614184. bioRxiv. 2024. PMID: 39386704 Free PMC article. Preprint.
-
Implications of Extended Inhibitory Neuron Development.Int J Mol Sci. 2021 May 12;22(10):5113. doi: 10.3390/ijms22105113. Int J Mol Sci. 2021. PMID: 34066025 Free PMC article. Review.
-
Understanding the heterogeneity of anxiety using a translational neuroscience approach.Cogn Affect Behav Neurosci. 2024 Apr;24(2):228-245. doi: 10.3758/s13415-024-01162-3. Epub 2024 Feb 14. Cogn Affect Behav Neurosci. 2024. PMID: 38356013 Free PMC article. Review.
-
Variation and convergence in the morpho-functional properties of the mammalian neocortex.Front Syst Neurosci. 2024 Jun 20;18:1413780. doi: 10.3389/fnsys.2024.1413780. eCollection 2024. Front Syst Neurosci. 2024. PMID: 38966330 Free PMC article. Review.
References
-
- Kumar S, Stecher G, Suleski M & Hedges SB TimeTree: a resource for timelines, timetrees, and divergence times. Mol. Biol. Evol 34, 1812–1819 (2017). - PubMed
-
- Northcutt RG & Kaas JH The emergence and evolution of mammalian neocortex. Trends Neurosci. 18, 373–379 (1995). - PubMed
-
- Laclef C & Métin C Conserved rules in embryonic development of cortical interneurons. Semin. Cell Dev. Biol 76, 86–100 (2018). - PubMed
-
- Katz PS & Harris-Warrick RM The evolution of neuronal circuits underlying species-specific behavior. Curr. Opin. Neurobiol 9, 628–633 (1999). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
