

Phase Separation of Disease-Associated SHP2 Mutants Underlies MAPK Hyperactivation
- PMID: 33002410
- PMCID: PMC7572904
- DOI: 10.1016/j.cell.2020.09.002
Phase Separation of Disease-Associated SHP2 Mutants Underlies MAPK Hyperactivation
Abstract
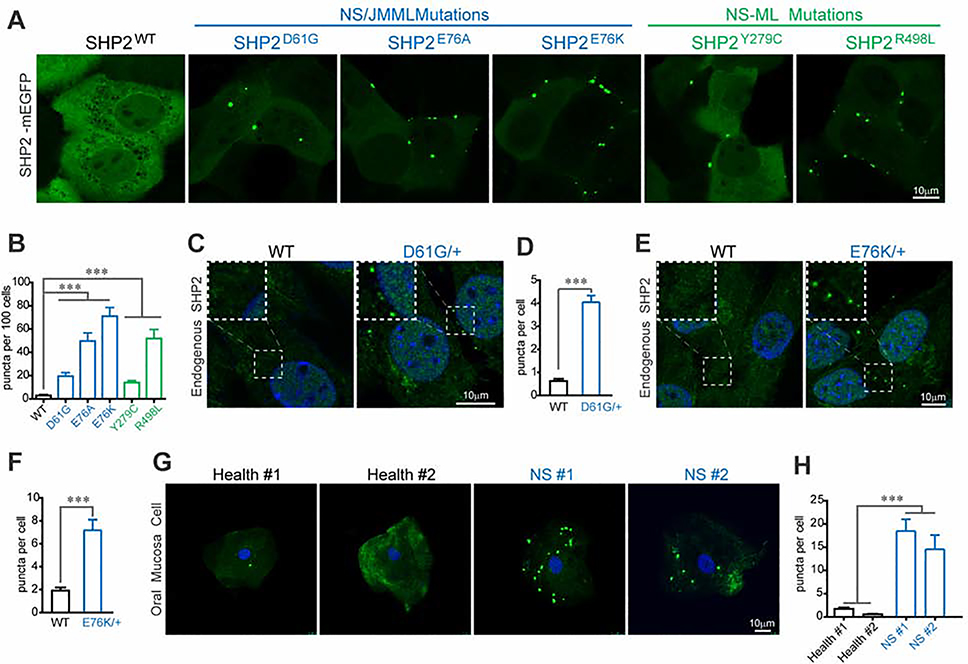
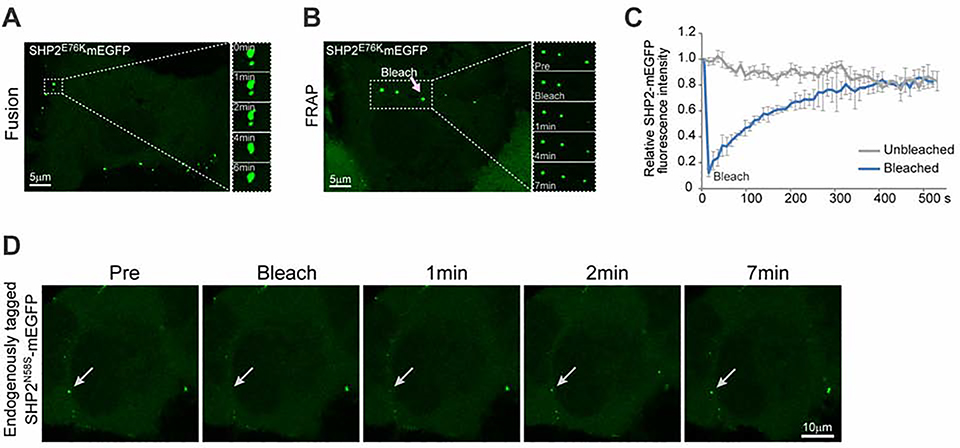
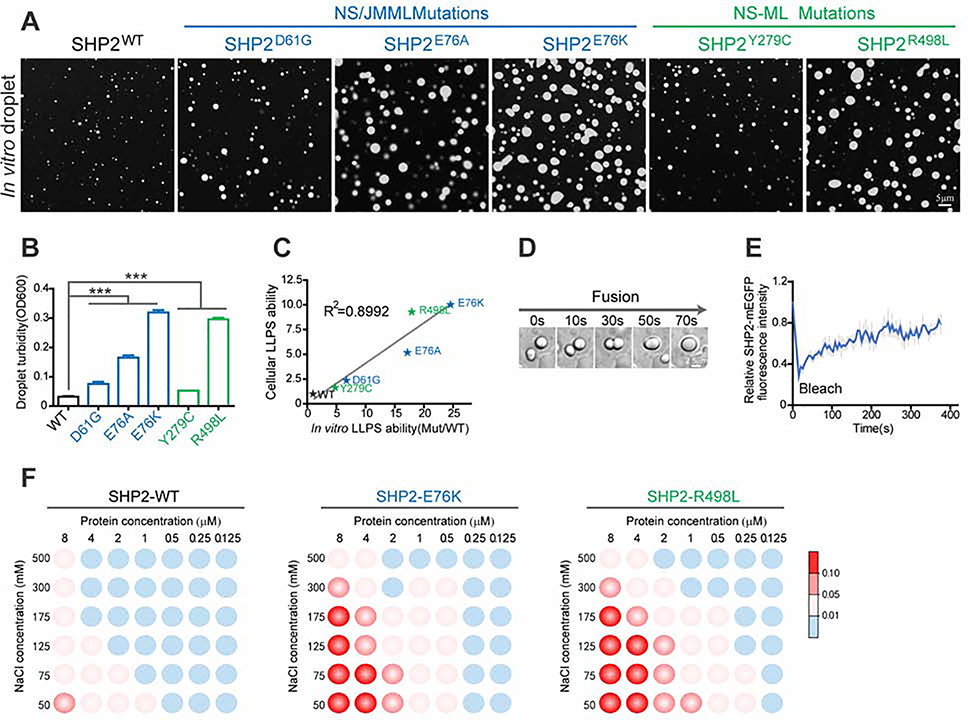
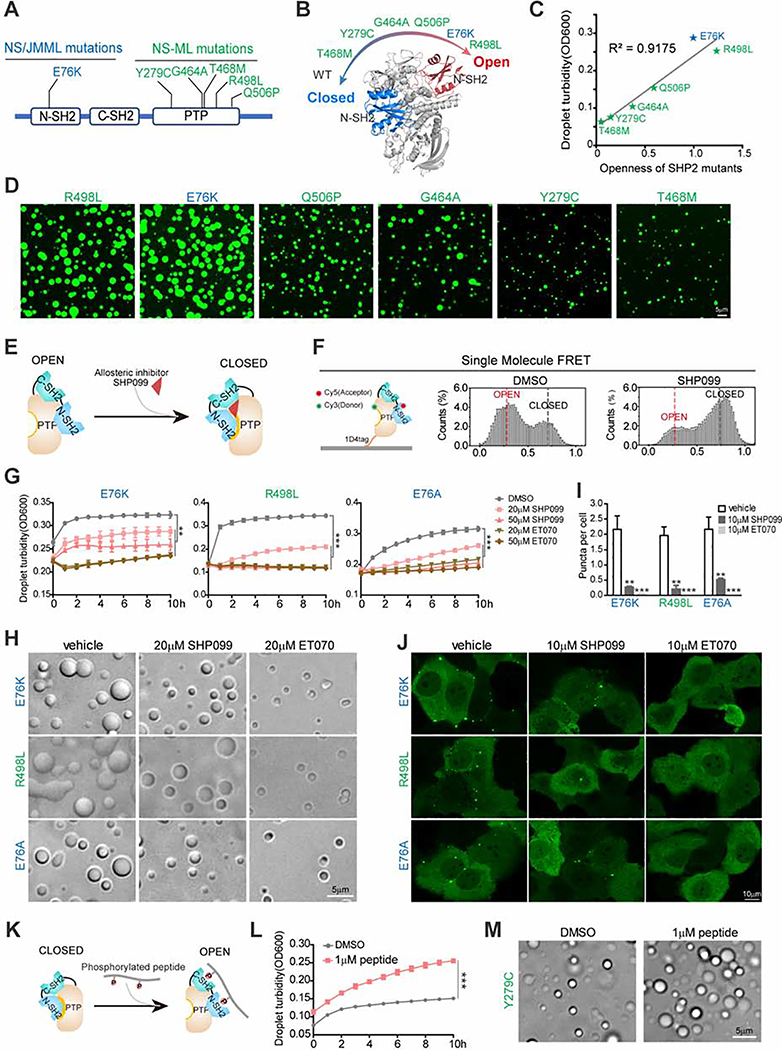
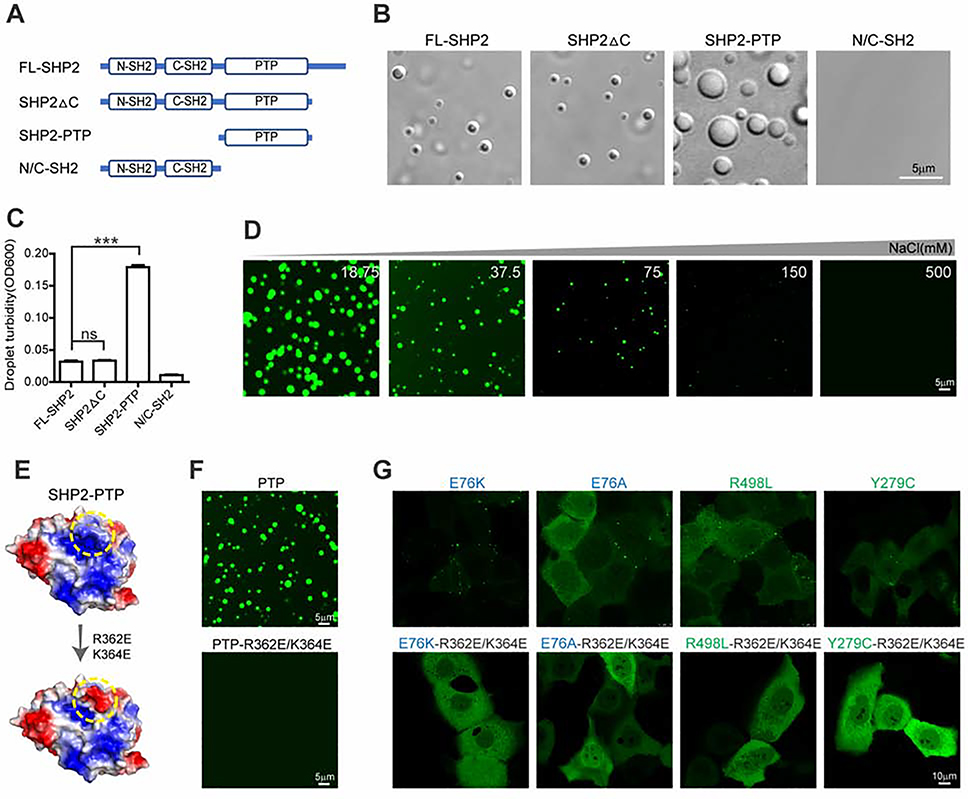
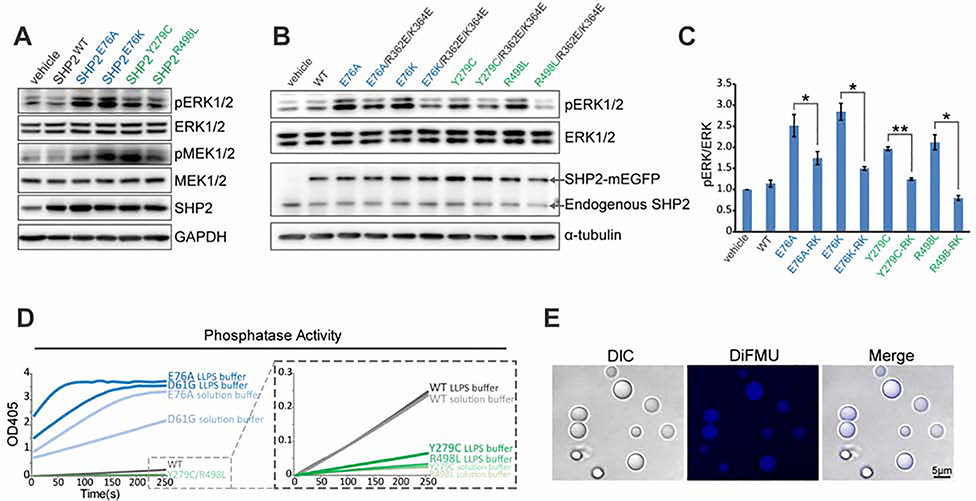
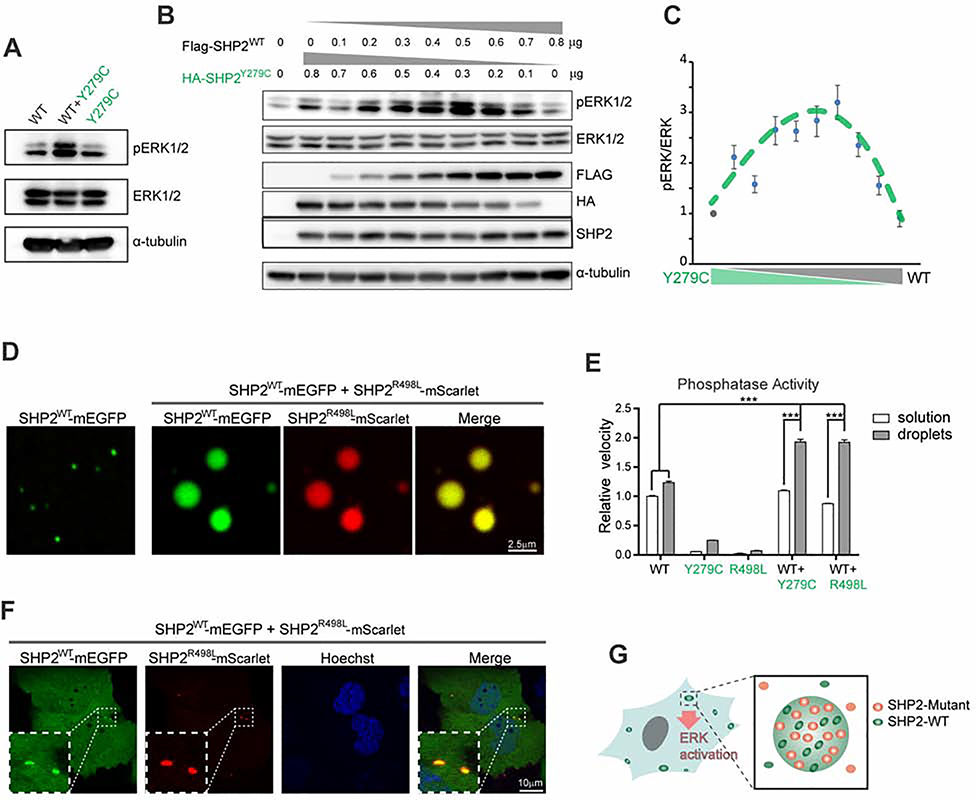
The non-receptor protein tyrosine phosphatase (PTP) SHP2, encoded by PTPN11, plays an essential role in RAS-mitogen-activated protein kinase (MAPK) signaling during normal development. It has been perplexing as to why both enzymatically activating and inactivating mutations in PTPN11 result in human developmental disorders with overlapping clinical manifestations. Here, we uncover a common liquid-liquid phase separation (LLPS) behavior shared by these disease-associated SHP2 mutants. SHP2 LLPS is mediated by the conserved well-folded PTP domain through multivalent electrostatic interactions and regulated by an intrinsic autoinhibitory mechanism through conformational changes. SHP2 allosteric inhibitors can attenuate LLPS of SHP2 mutants, which boosts SHP2 PTP activity. Moreover, disease-associated SHP2 mutants can recruit and activate wild-type (WT) SHP2 in LLPS to promote MAPK activation. These results not only suggest that LLPS serves as a gain-of-function mechanism involved in the pathogenesis of SHP2-associated human diseases but also provide evidence that PTP may be regulated by LLPS that can be therapeutically targeted.
Keywords: MAPK activation; Noonan syndrome; Noonan syndrome with multiple lentigines; PTPN11; SHP2; allosteric inhibitor; conformation change; disease-associated mutants; electrostatic interactions; liquid-liquid phase separation.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests J.Z. is a co-founder of Etern Biopharma Co. Ltd. and a member of its scientific advisory board. Q.Z. is a consultant to Etern Biopharma Co. Ltd. L.D. is an employee of Etern Biopharma Co. Ltd. H.C.S. is an employee of Roche. Etern Biopharma Co. Ltd. holds the patent WO-2020094018 that includes ET070 as one exemplified compound.
Figures
References
-
- Alberti S, and Dormann D (2019). Liquid–Liquid Phase Separation in Disease. Annu. Rev. Genet 53,3.1–3.24 - PubMed
-
- Araki T, Mohi MG, Ismat FA, Bronson RT, Williams IR, Kutok JL, Yang W, Pao LI, Gilliland DG, Epstein JA and Neel BG(2004). Mouse model of Noonan syndrome reveals cell type- and gene dosage- dependent effects of Ptpn11 mutation. Nat. Med 10, 849–857. - PubMed
-
- Böhmer F, Szedlacsek S, Tabernero L, Ostman A, and Den HJ (2013). Protein tyrosine phosphatase structure: function relationships in regulation and pathogenesis. Febs J. 280, 413–431. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
