

Major histocompatibility complex variation is similar in little brown bats before and after white-nose syndrome outbreak
- PMID: 33005361
- PMCID: PMC7520216
- DOI: 10.1002/ece3.6662
Major histocompatibility complex variation is similar in little brown bats before and after white-nose syndrome outbreak
Abstract
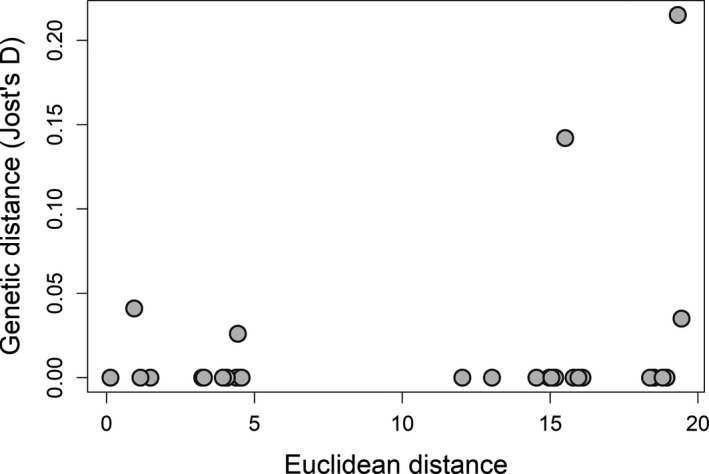
White-nose syndrome (WNS), caused by the fungal pathogen Pseudogymnoascus destructans (Pd), has driven alarming declines in North American hibernating bats, such as little brown bat (Myotis lucifugus). During hibernation, infected little brown bats are able to initiate anti-Pd immune responses, indicating pathogen-mediated selection on the major histocompatibility complex (MHC) genes. However, such immune responses may not be protective as they interrupt torpor, elevate energy costs, and potentially lead to higher mortality rates. To assess whether WNS drives selection on MHC genes, we compared the MHC DRB gene in little brown bats pre- (Wisconsin) and post- (Michigan, New York, Vermont, and Pennsylvania) WNS (detection spanning 2014-2015). We genotyped 131 individuals and found 45 nucleotide alleles (27 amino acid alleles) indicating a maximum of 3 loci (1-5 alleles per individual). We observed high allelic admixture and a lack of genetic differentiation both among sampling sites and between pre- and post-WNS populations, indicating no signal of selection on MHC genes. However, post-WNS populations exhibited decreased allelic richness, reflecting effects from bottleneck and drift following rapid population declines. We propose that mechanisms other than adaptive immunity are more likely driving current persistence of little brown bats in affected regions.
Keywords: Myotis lucifugus; North American bat; Pseudogymnoascus destructans; fungal disease; immunity; major histocompatibility complex.
© 2020 The Authors. Ecology and Evolution published by John Wiley & Sons Ltd.
Conflict of interest statement
No conflict of interest is declared by any of the authors of this manuscript.
Figures




Similar articles
-
Antibodies to Pseudogymnoascus destructans are not sufficient for protection against white-nose syndrome.Ecol Evol. 2015 Jun;5(11):2203-14. doi: 10.1002/ece3.1502. Epub 2015 May 11. Ecol Evol. 2015. PMID: 26078857 Free PMC article.
-
Experimental Infection of Tadarida brasiliensis with Pseudogymnoascus destructans, the Fungus That Causes White-Nose Syndrome.mSphere. 2018 Aug 29;3(4):e00250-18. doi: 10.1128/mSphere.00250-18. mSphere. 2018. PMID: 30158282 Free PMC article.
-
White-nose syndrome survivors do not exhibit frequent arousals associated with Pseudogymnoascus destructans infection.Front Zool. 2016 Mar 3;13:12. doi: 10.1186/s12983-016-0143-3. eCollection 2016. Front Zool. 2016. PMID: 26949407 Free PMC article.
-
COULD WHITE-NOSE SYNDROME MANIFEST DIFFERENTLY IN MYOTIS LUCIFUGUS IN WESTERN VERSUS EASTERN REGIONS OF NORTH AMERICA? A REVIEW OF FACTORS.J Wildl Dis. 2023 Jul 1;59(3):381-397. doi: 10.7589/JWD-D-22-00050. J Wildl Dis. 2023. PMID: 37270186 Review.
-
Investigating and managing the rapid emergence of white-nose syndrome, a novel, fatal, infectious disease of hibernating bats.Conserv Biol. 2011 Apr;25(2):223-31. doi: 10.1111/j.1523-1739.2010.01638.x. Epub 2011 Feb 1. Conserv Biol. 2011. PMID: 21284732 Review.
Cited by
-
Diversifying selection and climatic effects on major histocompatibility complex class II gene diversity in the greater horseshoe bat.Evol Appl. 2023 Jan 8;16(3):688-704. doi: 10.1111/eva.13528. eCollection 2023 Mar. Evol Appl. 2023. PMID: 36969140 Free PMC article.
-
Improved high-throughput MHC typing for non-model species using long-read sequencing.Mol Ecol Resour. 2022 Apr;22(3):862-876. doi: 10.1111/1755-0998.13511. Epub 2021 Oct 6. Mol Ecol Resour. 2022. PMID: 34551192 Free PMC article.
-
Selection and demography drive range-wide patterns of MHC-DRB variation in mule deer.BMC Ecol Evol. 2022 Apr 6;22(1):42. doi: 10.1186/s12862-022-01998-8. BMC Ecol Evol. 2022. PMID: 35387584 Free PMC article.
References
-
- Bataille, A. , Cashins, S. D. , Grogan, L. , Skerratt, L. F. , Hunter, D. , McFadden, M. , … Waldman, B. (2015). Susceptibility of amphibians to chytridiomycosis is associated with MHC class II conformation. Proceedings of the Royal Society B: Biological Sciences, 282(1805), 20143127 10.1098/rspb.2014.3127 - DOI - PMC - PubMed
-
- Baucom, R. S. , & de Roode, J. C. (2011). Ecological immunology and tolerance in plants and animals. Functional Ecology, 25(1), 18–28. 10.1111/j.1365-2435.2010.01742.x - DOI
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
