

Therapeutic potential of extracellular vesicle-associated long noncoding RNA
- PMID: 33005738
- PMCID: PMC7510462
- DOI: 10.1002/btm2.10172
Therapeutic potential of extracellular vesicle-associated long noncoding RNA
Abstract
Both extracellular vesicles (EVs) and long noncoding RNAs (lncRNAs) have been increasingly investigated as biomarkers, pathophysiological mediators, and potential therapeutics. While these two entities have often been studied separately, there are increasing reports of EV-associated lncRNA activity in processes such as oncogenesis as well as tissue repair and regeneration. Given the powerful nature and emerging translational impact of other noncoding RNAs such as microRNA (miRNA) and small interfering RNA, lncRNA therapeutics may represent a new frontier. While EVs are natural vehicles that transport and protect lncRNAs physiologically, they can also be engineered for enhanced cargo loading and therapeutic properties. In this review, we will summarize the activity of lncRNAs relevant to both tissue repair and cancer treatment and discuss the role of EVs in enabling the potential of lncRNA therapeutics.
Keywords: exosomes; extracellular vesicles; lncRNA; microparticles; microvesicles.
© 2020 The Authors. Bioengineering & Translational Medicine published by Wiley Periodicals LLC on behalf of American Institute of Chemical Engineers.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
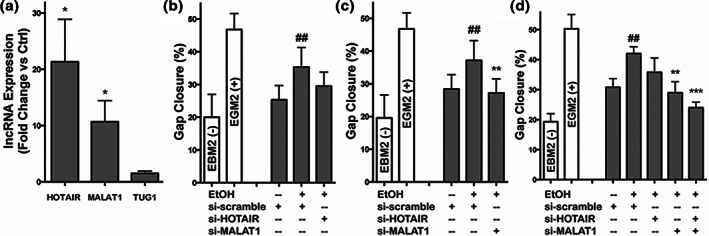

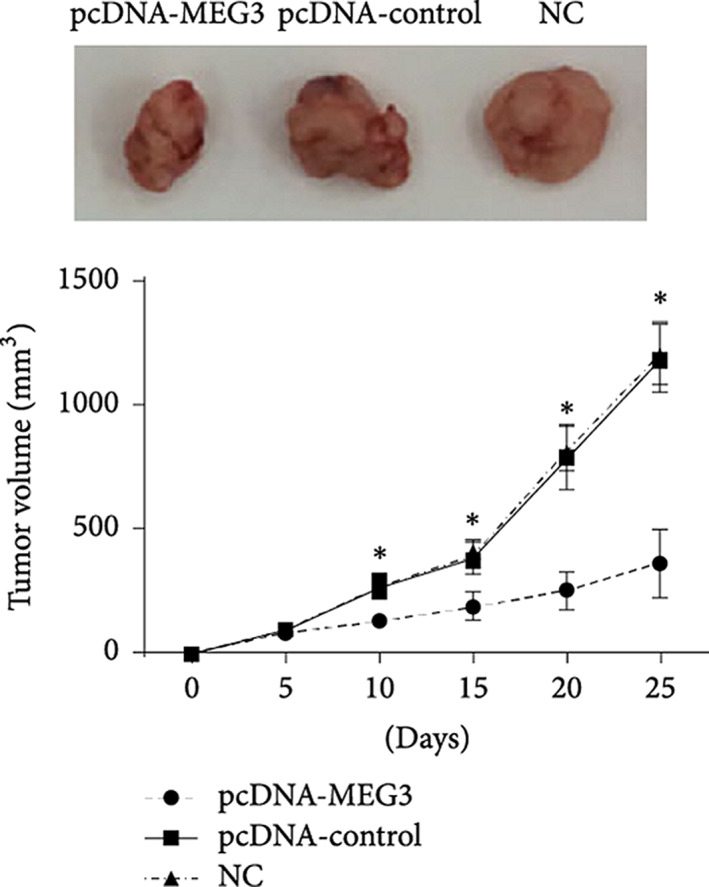

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
