

Phase Separation in Germ Cells and Development
- PMID: 33007213
- PMCID: PMC7572804
- DOI: 10.1016/j.devcel.2020.09.004
Phase Separation in Germ Cells and Development
Abstract
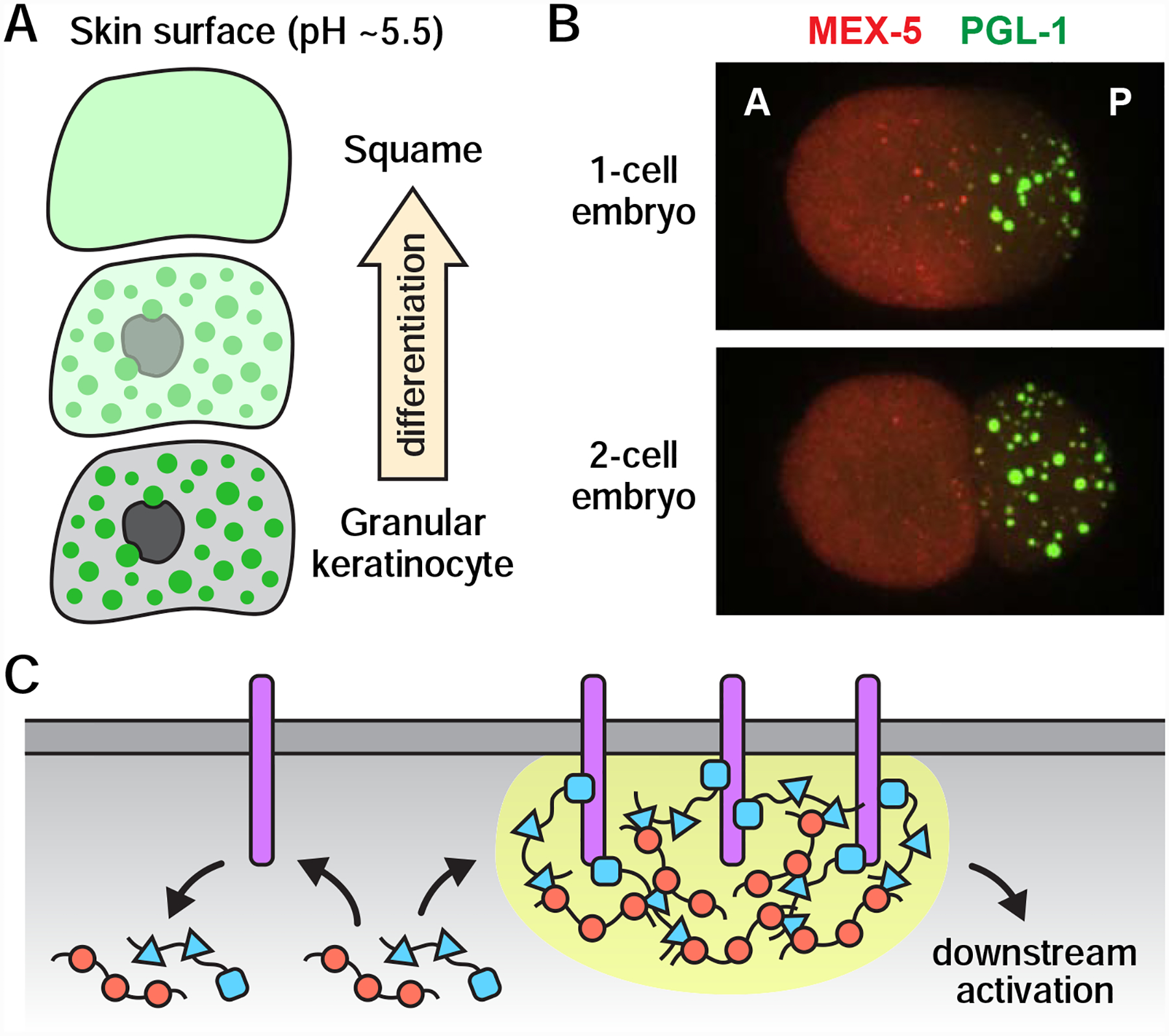
The animal germline is an immortal cell lineage that gives rise to eggs and/or sperm each generation. Fusion of an egg and sperm, or fertilization, sets off a cascade of developmental events capable of producing an array of different cell types and body plans. How germ cells develop, function, and eventually give rise to entirely new organisms is an important question in biology. A growing body of evidence suggests that phase separation events likely play a significant and multifaceted role in germ cells and development. Here, we discuss the organization, dynamics, and potential functions of phase-separated compartments in germ cells and examine the various ways in which phase separation might contribute to the development of multicellular organisms.
Keywords: biomolecular condensates; development; germ granules; phase separation.
Copyright © 2020 Elsevier Inc. All rights reserved.
Figures




References
-
- Alberti S (2017) ‘The wisdom of crowds: regulating cell function through condensed states of living matter’, Journal of cell science, 130(17), pp. 2789–2796. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
