

Structural diversity and phylogenetic distribution of valyl tRNA-like structures in viruses
- PMID: 33008837
- PMCID: PMC7749636
- DOI: 10.1261/rna.076968.120
Structural diversity and phylogenetic distribution of valyl tRNA-like structures in viruses
Abstract
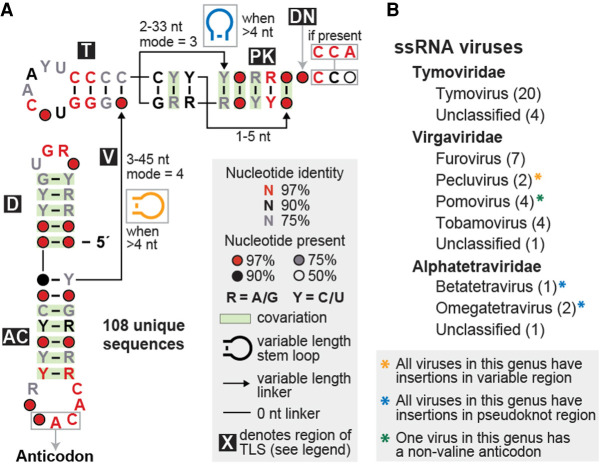
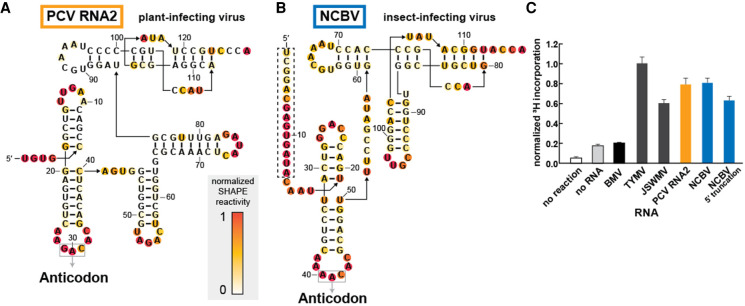
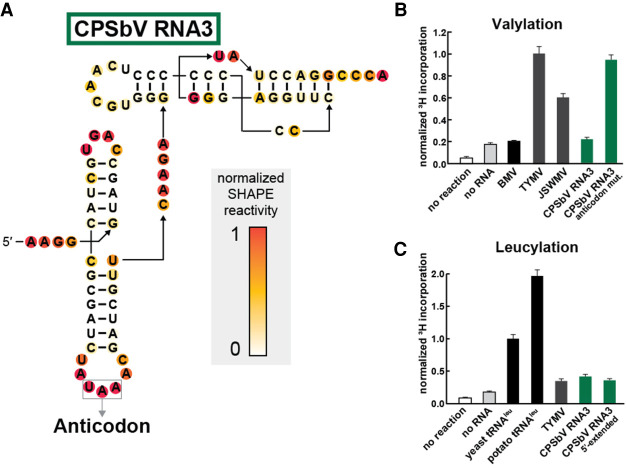
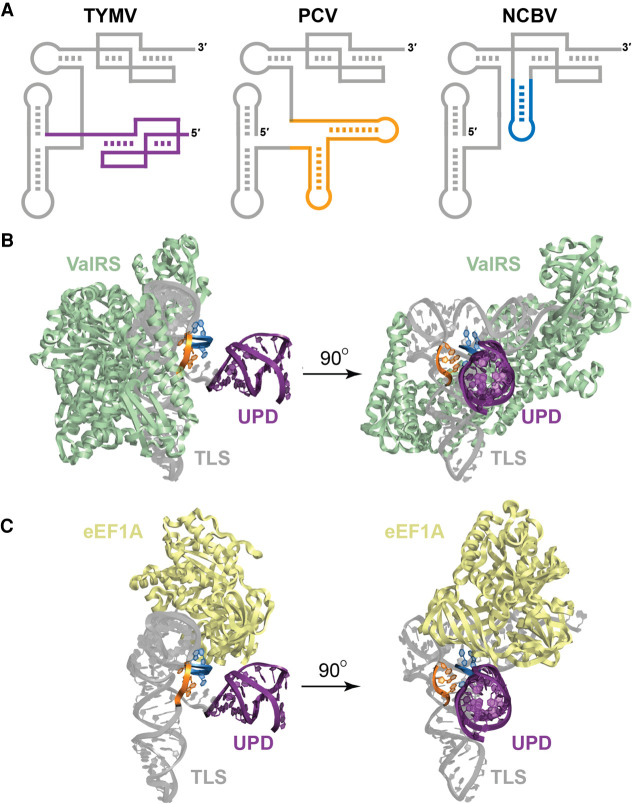
Viruses commonly use specifically folded RNA elements that interact with both host and viral proteins to perform functions important for diverse viral processes. Examples are found at the 3' termini of certain positive-sense ssRNA virus genomes where they partially mimic tRNAs, including being aminoacylated by host cell enzymes. Valine-accepting tRNA-like structures (TLSVal) are an example that share some clear homology with canonical tRNAs but have several important structural differences. Although many examples of TLSVal have been identified, we lacked a full understanding of their structural diversity and phylogenetic distribution. To address this, we undertook an in-depth bioinformatic and biochemical investigation of these RNAs, guided by recent high-resolution structures of a TLSVal We cataloged many new examples in plant-infecting viruses but also in unrelated insect-specific viruses. Using biochemical and structural approaches, we verified the secondary structure of representative TLSVal substrates and tested their ability to be valylated, confirming previous observations of structural heterogeneity within this class. In a few cases, large stem-loop structures are inserted within variable regions located in an area of the TLS distal to known host cell factor binding sites. In addition, we identified one virus whose TLS has switched its anticodon away from valine, causing a loss of valylation activity; the implications of this remain unclear. These results refine our understanding of the structural and functional mechanistic details of tRNA mimicry and how this may be used in viral infection.
Keywords: RNA structure; aminoacylation; tRNA mimicry; viral RNA.
© 2021 Sherlock et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources