

Phylogenomic analysis sheds light on the evolutionary pathways towards acoustic communication in Orthoptera
- PMID: 33009390
- PMCID: PMC7532154
- DOI: 10.1038/s41467-020-18739-4
Phylogenomic analysis sheds light on the evolutionary pathways towards acoustic communication in Orthoptera
Erratum in
-
Author Correction: Phylogenomic analysis sheds light on the evolutionary pathways towards acoustic communication in Orthoptera.Nat Commun. 2020 Nov 3;11(1):5674. doi: 10.1038/s41467-020-19626-8. Nat Commun. 2020. PMID: 33144590 Free PMC article.
Abstract
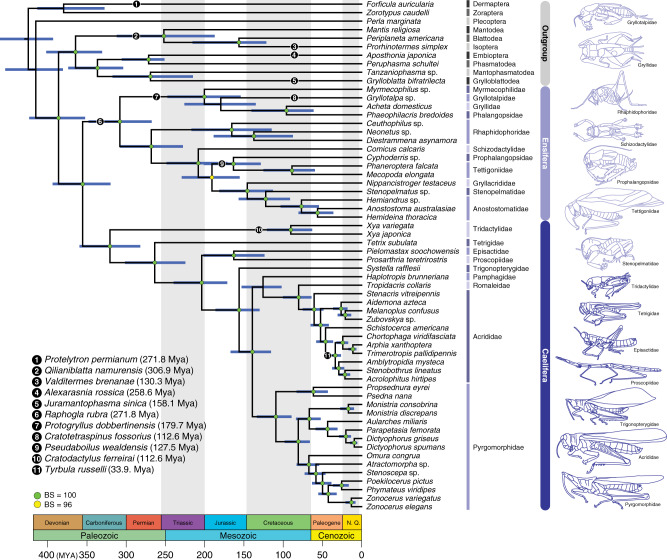
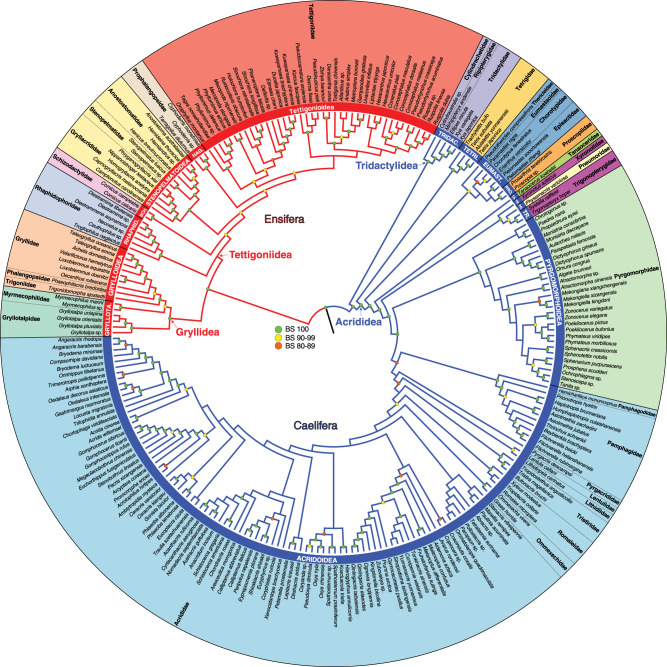
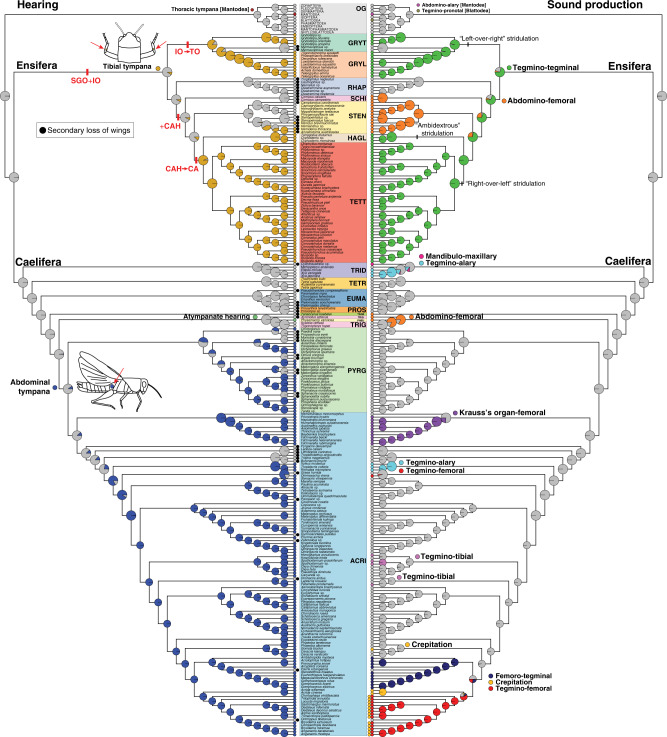
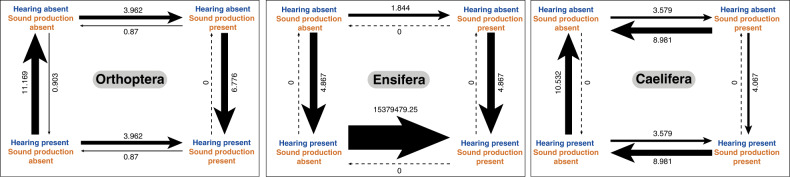
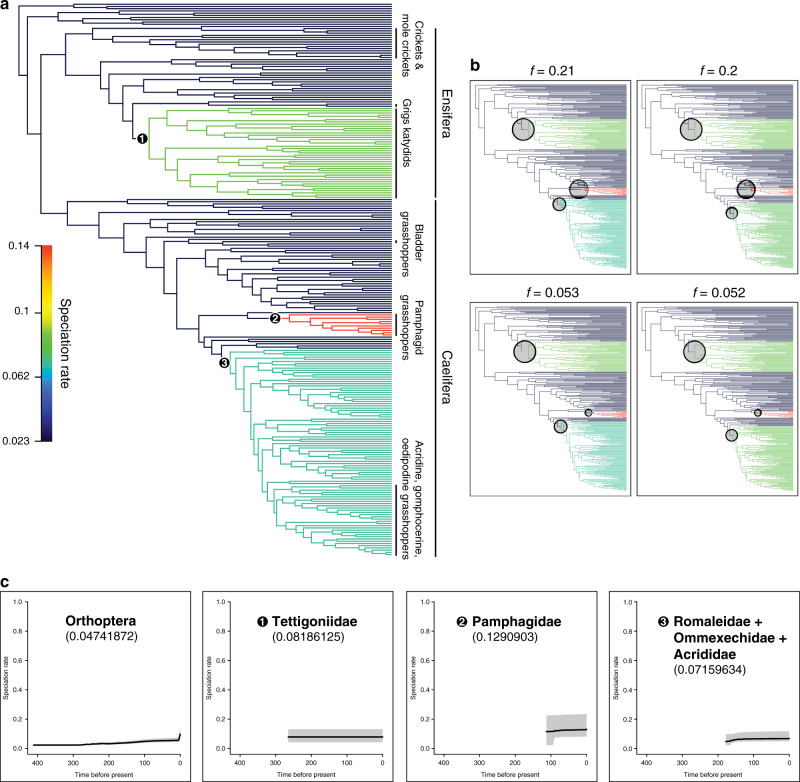
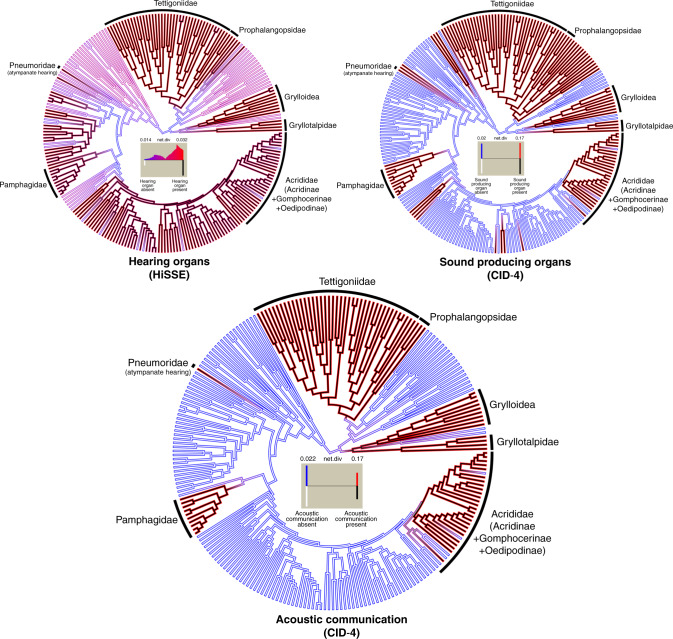
Acoustic communication is enabled by the evolution of specialised hearing and sound producing organs. In this study, we performed a large-scale macroevolutionary study to understand how both hearing and sound production evolved and affected diversification in the insect order Orthoptera, which includes many familiar singing insects, such as crickets, katydids, and grasshoppers. Using phylogenomic data, we firmly establish phylogenetic relationships among the major lineages and divergence time estimates within Orthoptera, as well as the lineage-specific and dynamic patterns of evolution for hearing and sound producing organs. In the suborder Ensifera, we infer that forewing-based stridulation and tibial tympanal ears co-evolved, but in the suborder Caelifera, abdominal tympanal ears first evolved in a non-sexual context, and later co-opted for sexual signalling when sound producing organs evolved. However, we find little evidence that the evolution of hearing and sound producing organs increased diversification rates in those lineages with known acoustic communication.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Brumm, H. Animal Communication and Noise (Springer, 2013).
-
- Hedwig, B. Insect Hearing and Acoustic Communication (Springer, 2014).
-
- Ewing, A. W. Arthropod Bioacoustics: Neurobiology and Behaviour (Cornell Univ. Press, 1989).
-
- Ladich F, Winkler H. Acoustic communication in terrestrial and aquatic vertebrates. J. Exp. Biol. 2017;220:2306–2317. - PubMed
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
