

The Genetic Mechanisms Driving Diversification of the KIR Gene Cluster in Primates
- PMID: 33013938
- PMCID: PMC7516082
- DOI: 10.3389/fimmu.2020.582804
The Genetic Mechanisms Driving Diversification of the KIR Gene Cluster in Primates
Abstract
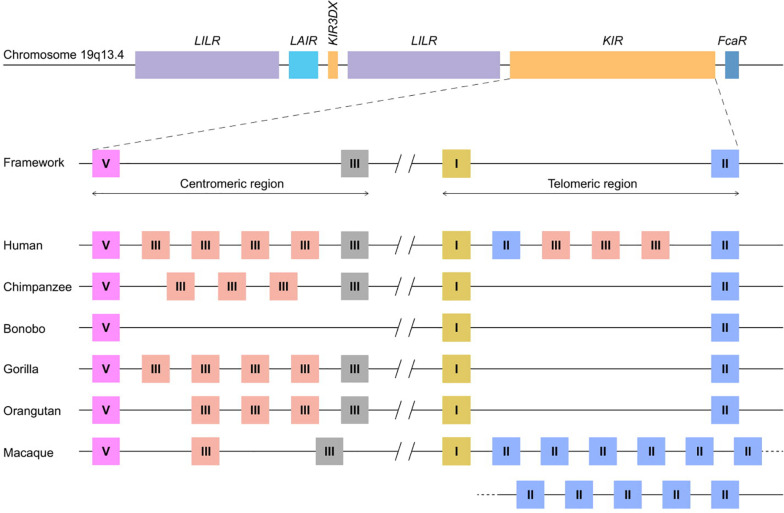
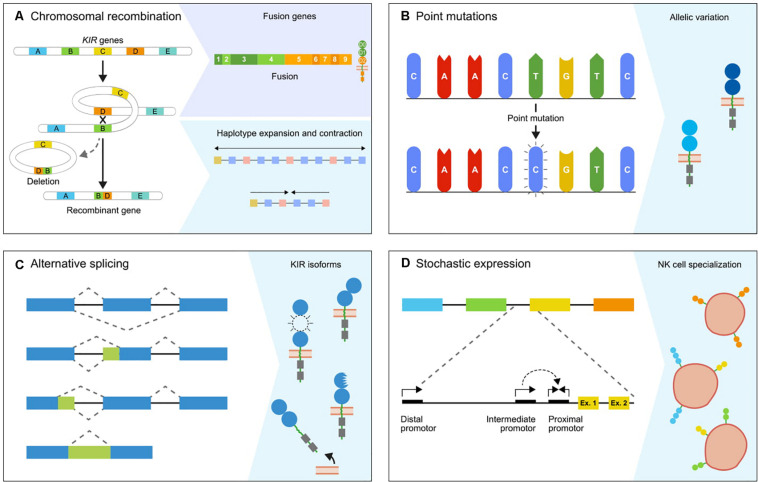
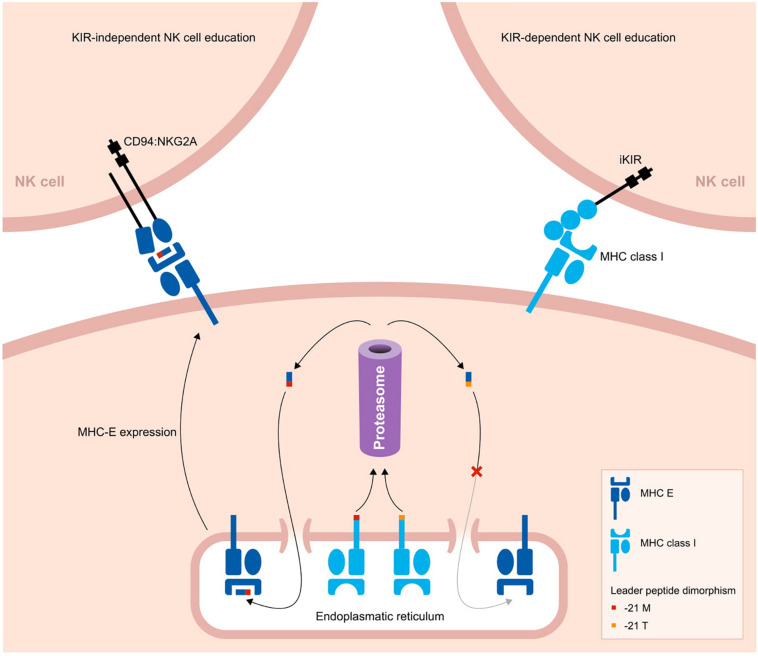
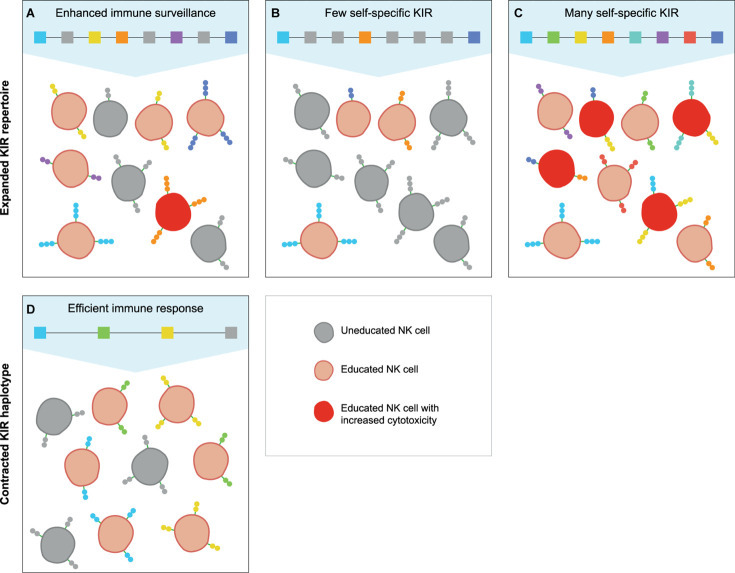
The activity and function of natural killer (NK) cells are modulated through the interactions of multiple receptor families, of which some recognize MHC class I molecules. The high level of MHC class I polymorphism requires their ligands either to interact with conserved epitopes, as is utilized by the NKG2A receptor family, or to co-evolve with the MHC class I allelic variation, which task is taken up by the killer cell immunoglobulin-like receptor (KIR) family. Multiple molecular mechanisms are responsible for the diversification of the KIR gene system, and include abundant chromosomal recombination, high mutation rates, alternative splicing, and variegated expression. The combination of these genetic mechanisms generates a compound array of diversity as is reflected by the contraction and expansion of KIR haplotypes, frequent birth of fusion genes, allelic polymorphism, structurally distinct isoforms, and variegated expression, which is in contrast to the mainly allelic nature of MHC class I polymorphism in humans. A comparison of the thoroughly studied human and macaque KIR gene repertoires demonstrates a similar evolutionarily conserved toolbox, through which selective forces drove and maintained the diversified nature of the KIR gene cluster. This hypothesis is further supported by the comparative genetics of KIR haplotypes and genes in other primate species. The complex nature of the KIR gene system has an impact upon the education, activity, and function of NK cells in coherence with an individual's MHC class I repertoire and pathogenic encounters. Although selection operates on an individual, the continuous diversification of the KIR gene system in primates might protect populations against evolving pathogens.
Keywords: KIR; Killer cell immunoglobin-like receptor; NK cell; NK cell education; human; macaque; non-human primates.
Copyright © 2020 Bruijnesteijn, de Groot and Bontrop.
Figures
References
-
- Vivier E, Tomasello E, Baratin M, Walzer T, Ugolini S. Functions of natural killer cells. Nat Immunol. (2008) 9:503–10. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials