

The Overlapping and Distinct Roles of HAM Family Genes in Arabidopsis Shoot Meristems
- PMID: 33013964
- PMCID: PMC7498855
- DOI: 10.3389/fpls.2020.541968
The Overlapping and Distinct Roles of HAM Family Genes in Arabidopsis Shoot Meristems
Abstract
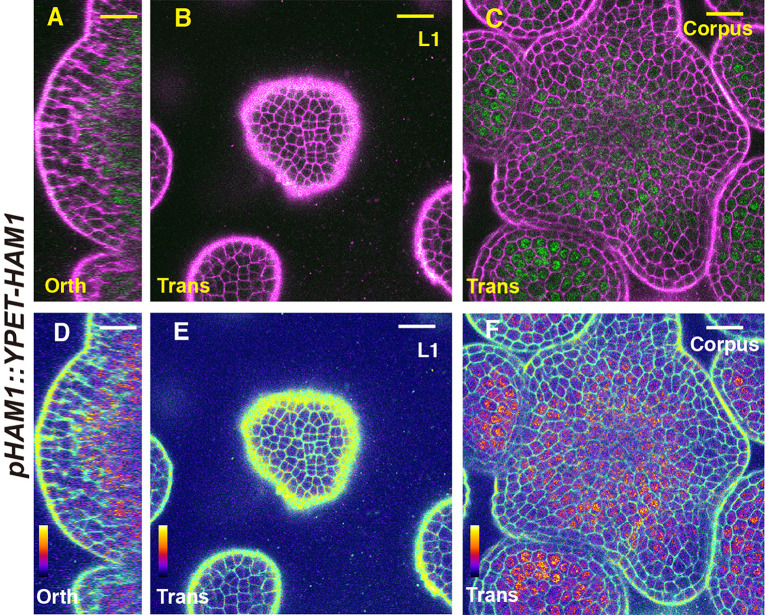
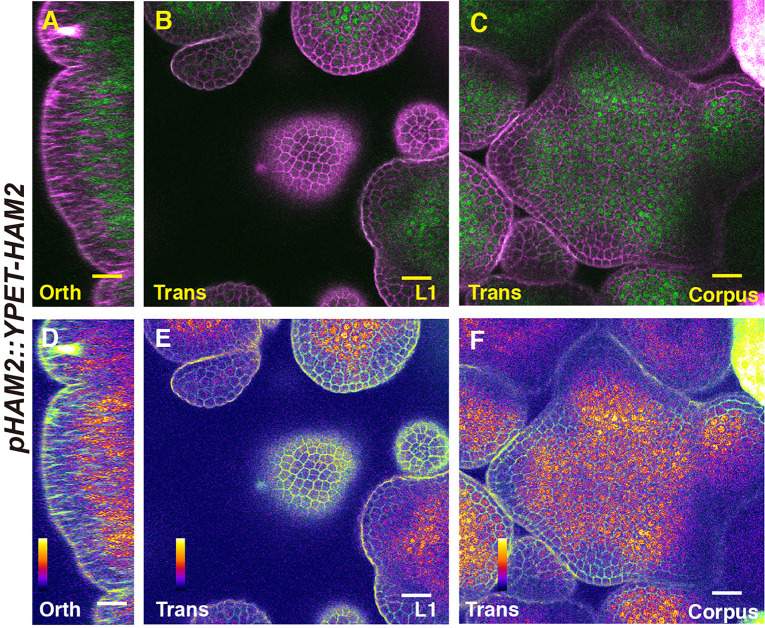
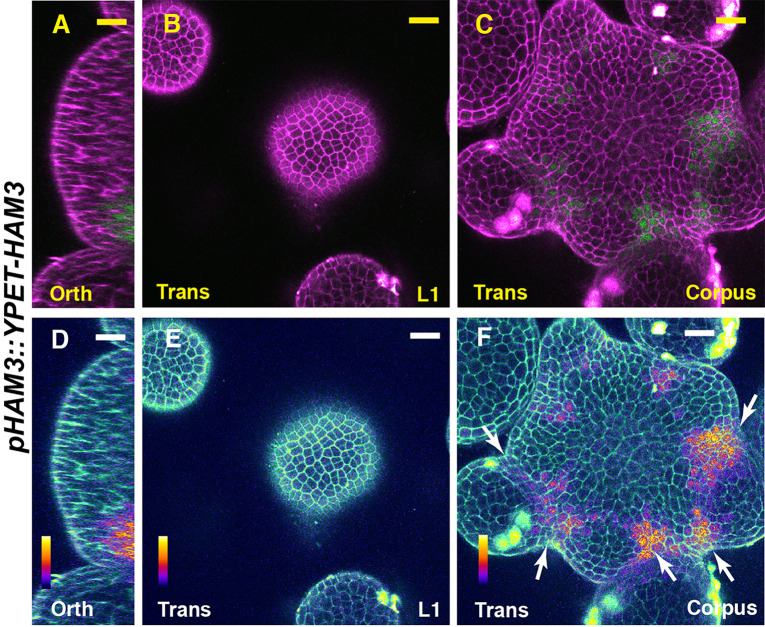
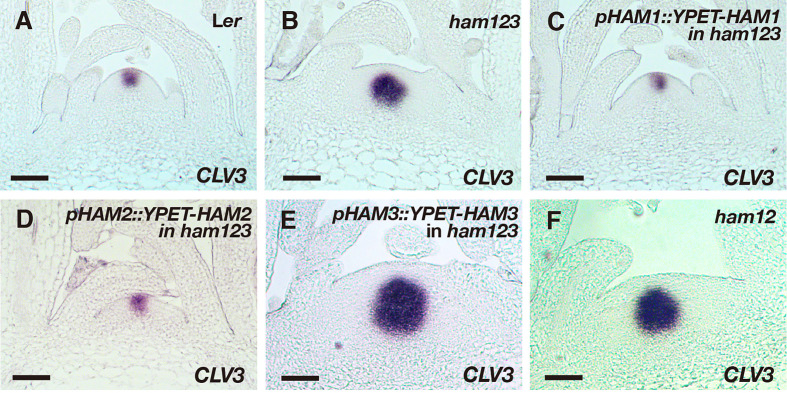
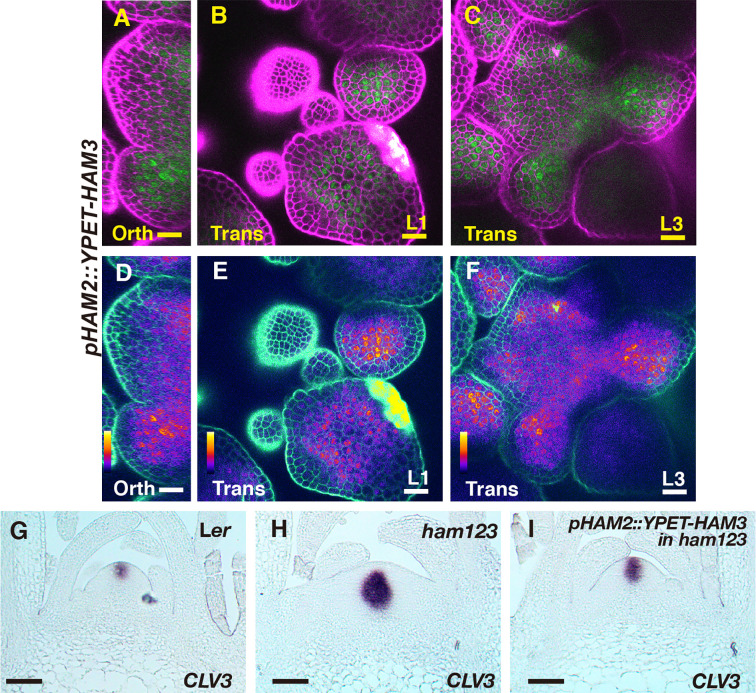
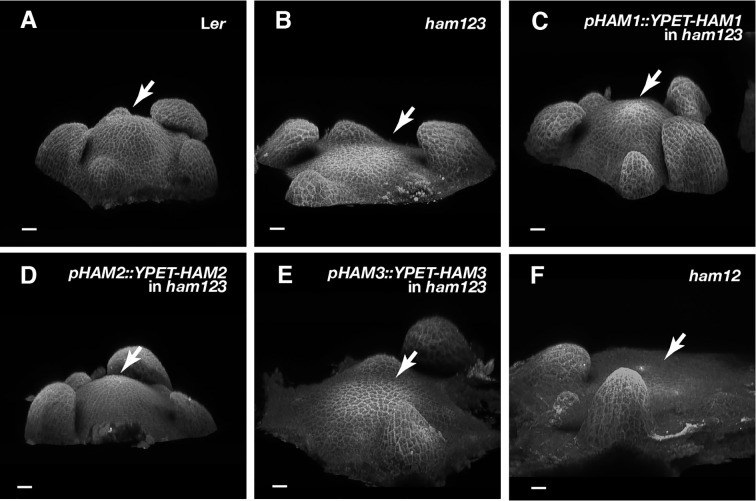
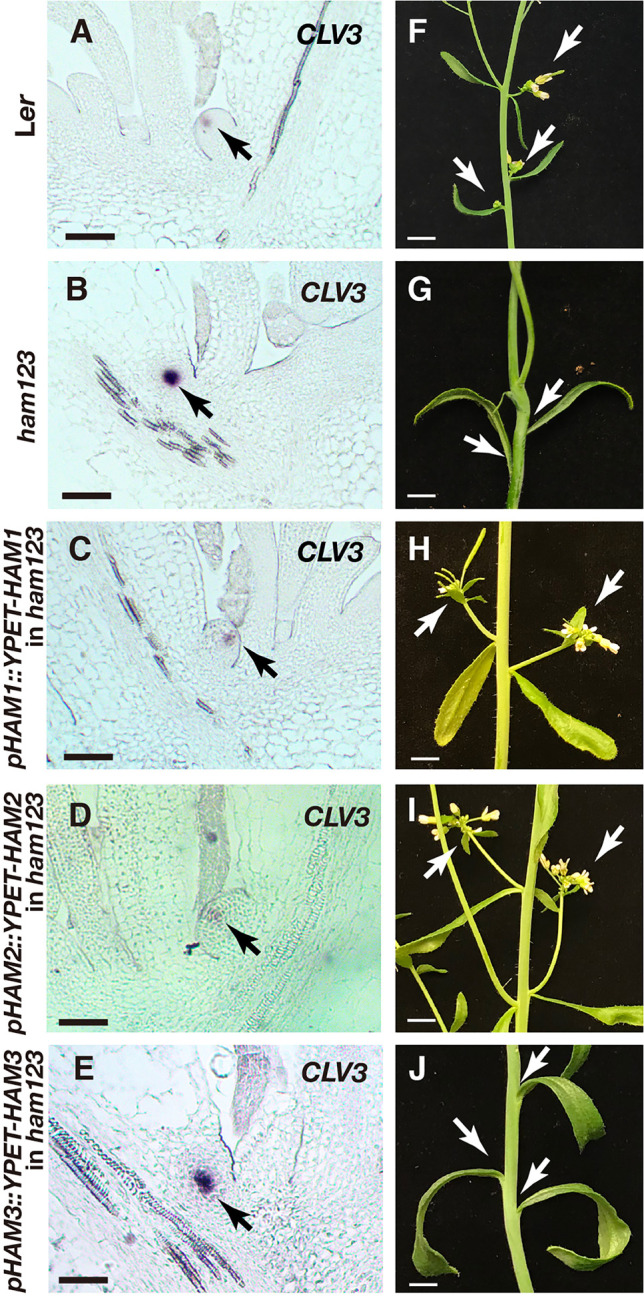
In Arabidopsis shoot apical meristems (SAMs), a well-characterized regulatory loop between WUSCHEL (WUS) and CLAVATA3 (CLV3) maintains stem cell homeostasis by regulating the balance between cell proliferation and cell differentiation. WUS proteins, translated in deep cell layers, move into the overlaying stem cells to activate CLV3. The secreted peptide CLV3 then regulates WUS levels through a ligand-receptor mediated signaling cascade. CLV3 is specifically expressed in the stem cells and repressed in the deep cell layers despite presence of the WUS activator, forming an apical-basal polarity along the axis of the SAM. Previously, we proposed and validated a hypothesis that the HAIRY MERISTEM (HAM) family genes regulate this polarity, keeping the expression of CLV3 off in interior cells of the SAM. However, the specific role of each individual member of the HAM family in this process remains to be elucidated. Combining live imaging and molecular genetics, we have dissected the conserved and distinct functions of different HAM family members in control of CLV3 patterning in the SAMs and in the de novo shoot stem cell niches as well.
Keywords: Arabidopsis; HAIRY MERISTEM; confocal imaging; shoot apical meristems; shoot development; stem cells.
Copyright © 2020 Han, Geng, Guo, Yan, Meyerowitz, Liu and Zhou.
Figures
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases