

Installation of C4 photosynthetic pathway enzymes in rice using a single construct
- PMID: 33016576
- PMCID: PMC7955876
- DOI: 10.1111/pbi.13487
Installation of C4 photosynthetic pathway enzymes in rice using a single construct
Abstract
Introduction of a C4 photosynthetic mechanism into C3 crops offers an opportunity to improve photosynthetic efficiency, biomass and yield in addition to potentially improving nitrogen and water use efficiency. To create a two-cell metabolic prototype for an NADP-malic enzyme type C4 rice, we transformed Oryza sativa spp. japonica cultivar Kitaake with a single construct containing the coding regions of carbonic anhydrase, phosphoenolpyruvate (PEP) carboxylase, NADP-malate dehydrogenase, pyruvate orthophosphate dikinase and NADP-malic enzyme from Zea mays, driven by cell-preferential promoters. Gene expression, protein accumulation and enzyme activity were confirmed for all five transgenes, and intercellular localization of proteins was analysed. 13 CO2 labelling demonstrated a 10-fold increase in flux though PEP carboxylase, exceeding the increase in measured in vitro enzyme activity, and estimated to be about 2% of the maize photosynthetic flux. Flux from malate via pyruvate to PEP remained low, commensurate with the low NADP-malic enzyme activity observed in the transgenic lines. Physiological perturbations were minor and RNA sequencing revealed no substantive effects of transgene expression on other endogenous rice transcripts associated with photosynthesis. These results provide promise that, with enhanced levels of the C4 proteins introduced thus far, a functional C4 pathway is achievable in rice.
Keywords: C4 photosynthesis; metabolic engineering; rice.
© 2020 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Conflict of interest statement
Authors declare no competing interests.
Figures


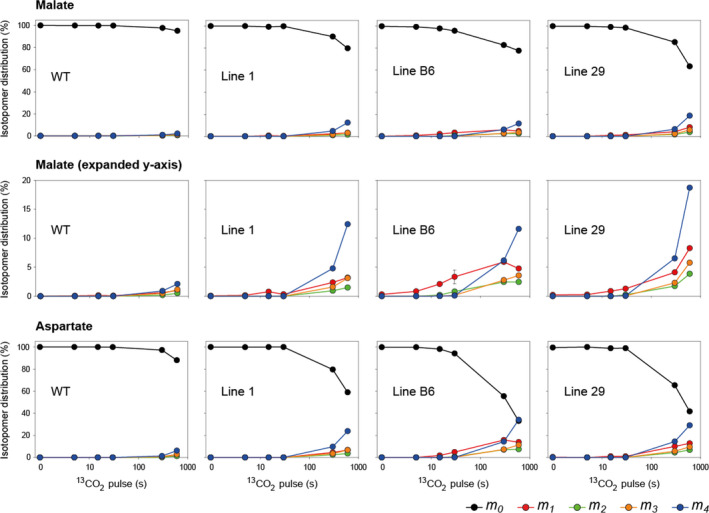
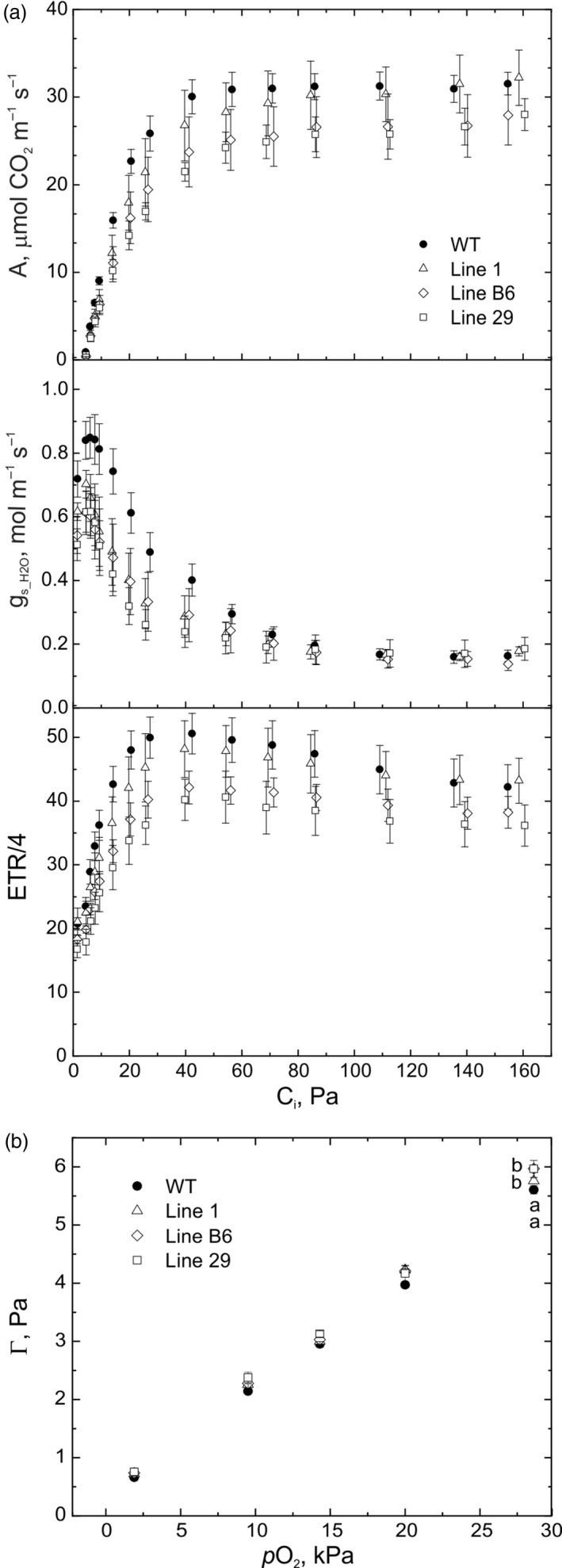
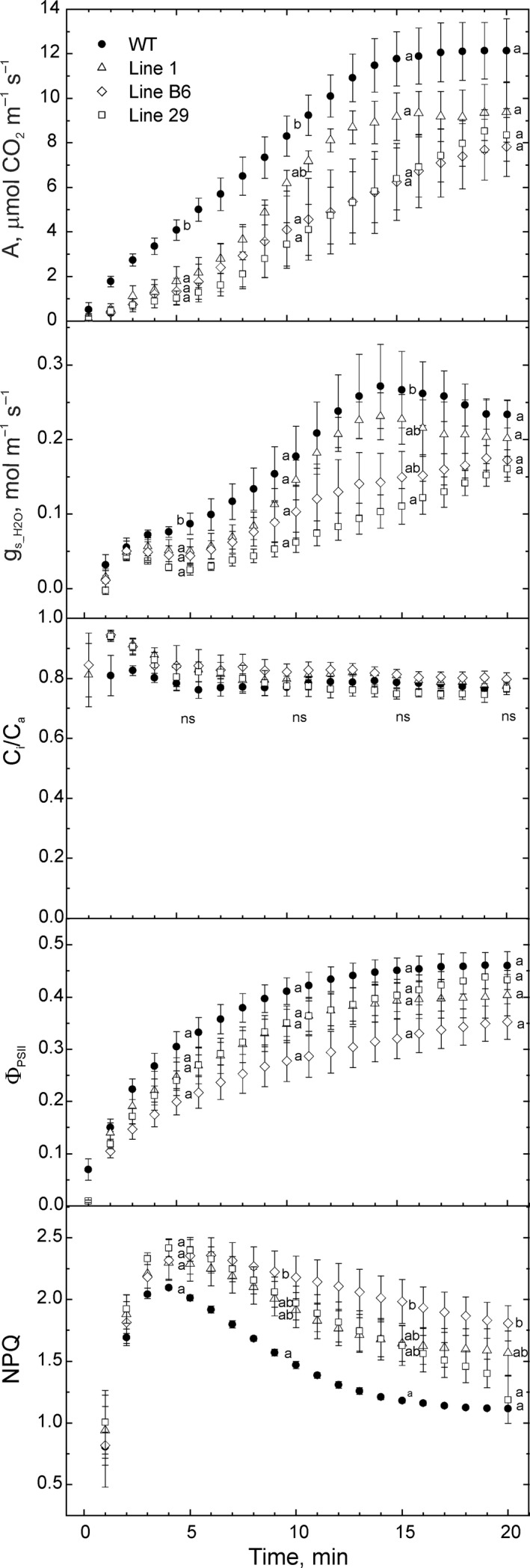
References
-
- Ashton, A.R. , Burnell, J.N. , Furbank, R.T. , Jenkins, C.L. and Hatch, M.D. (1990) Enzymes of C4 photosynthesis. In Methods in plant biochemistry ( Lea, P. , ed.), pp. 39–72. London: Academic press.
-
- Ashton, A.R. and Hatch, M.D. (1983) Regulation of C4 photosynthesis: Regulation of activation and inactivation of NADP‐malate dehydrogenase by NADP and NADPH. Arch. Biochem. Biophys. 227, 416–424. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
