

Microtubule affinity-regulating kinase 4 with an Alzheimer's disease-related mutation promotes tau accumulation and exacerbates neurodegeneration
- PMID: 33020179
- PMCID: PMC7863894
- DOI: 10.1074/jbc.RA120.014420
Microtubule affinity-regulating kinase 4 with an Alzheimer's disease-related mutation promotes tau accumulation and exacerbates neurodegeneration
Abstract
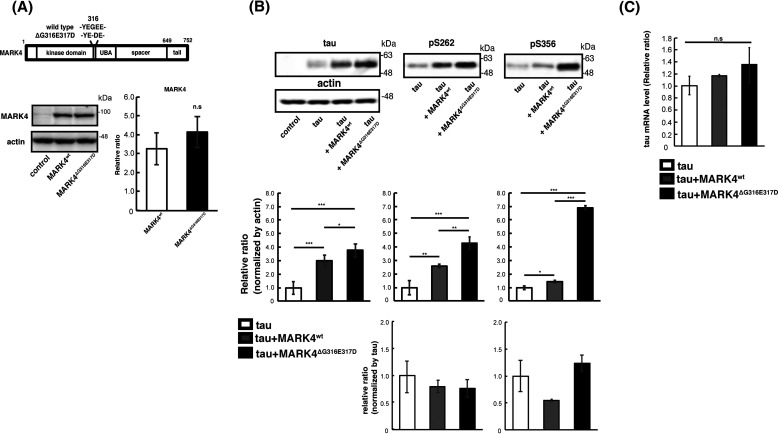
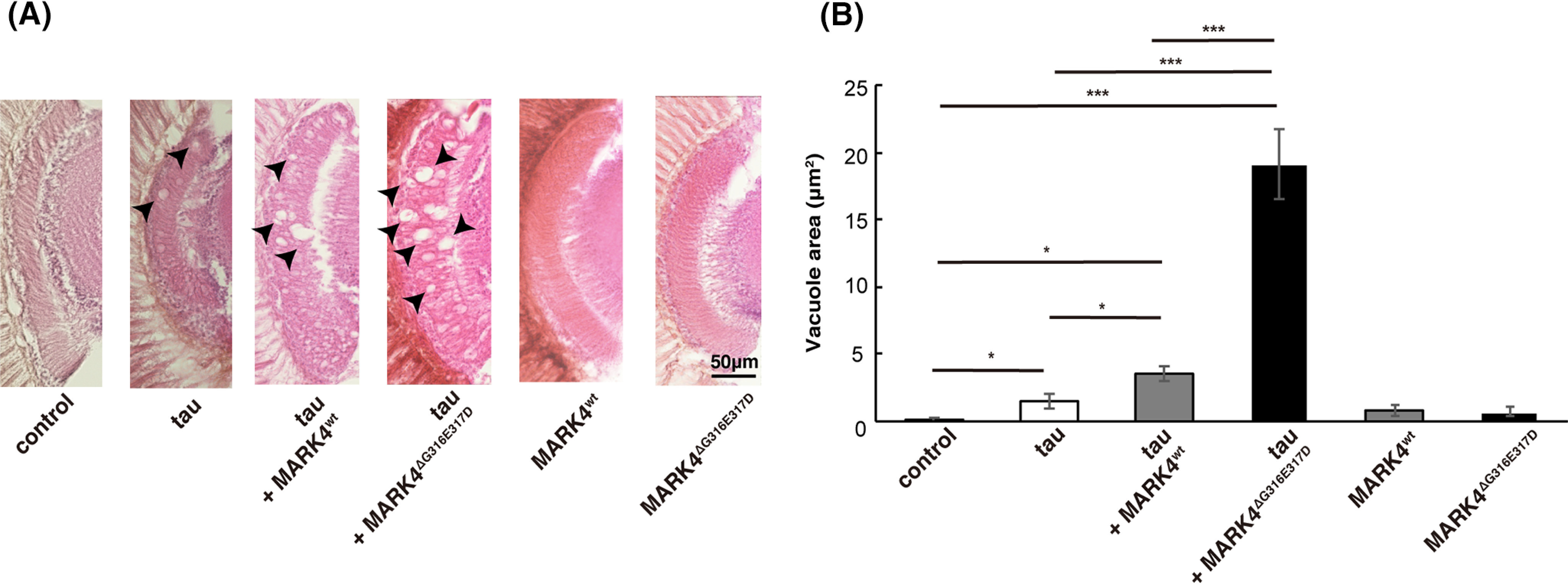
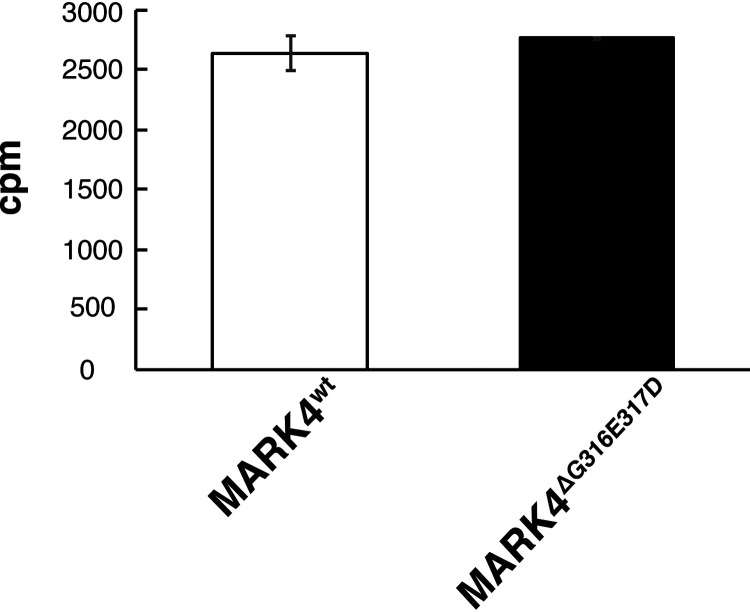
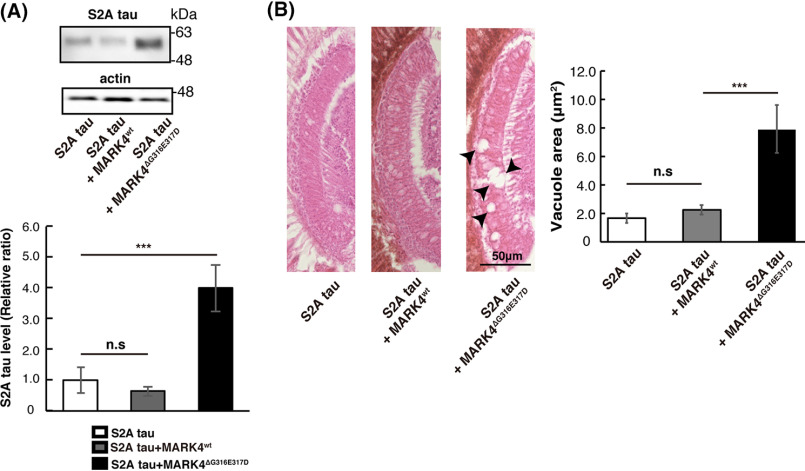
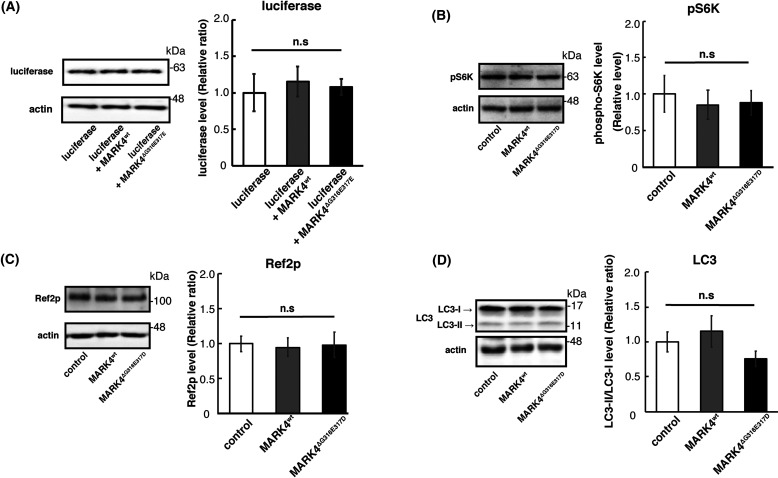
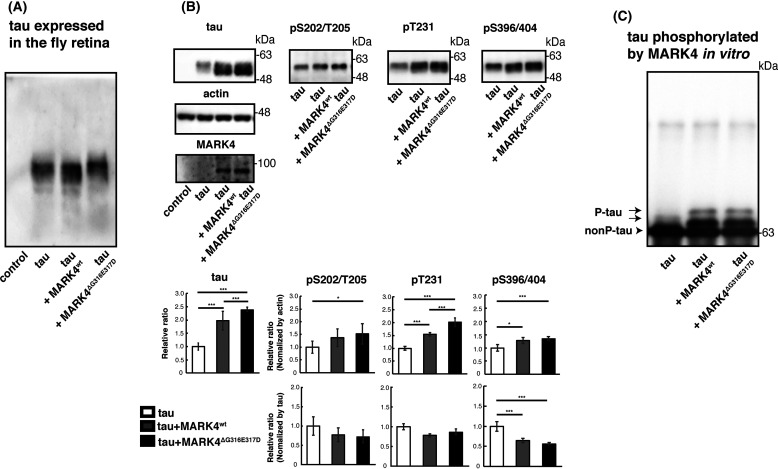
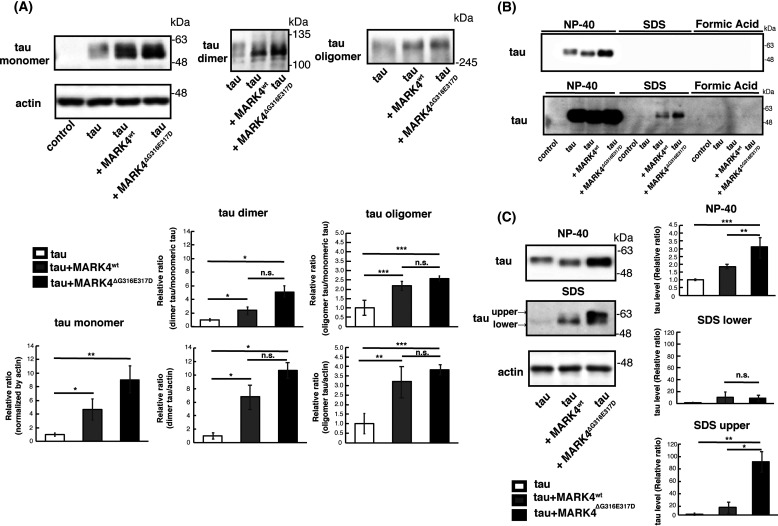
Accumulation of the microtubule-associated protein tau is associated with Alzheimer's disease (AD). In AD brain, tau is abnormally phosphorylated at many sites, and phosphorylation at Ser-262 and Ser-356 plays critical roles in tau accumulation and toxicity. Microtubule affinity-regulating kinase 4 (MARK4) phosphorylates tau at those sites, and a double de novo mutation in the linker region of MARK4, ΔG316E317D, is associated with an elevated risk of AD. However, it remains unclear how this mutation affects phosphorylation, aggregation, and accumulation of tau and tau-induced neurodegeneration. Here, we report that MARK4ΔG316E317D increases the abundance of highly phosphorylated, insoluble tau species and exacerbates neurodegeneration via Ser-262/356-dependent and -independent mechanisms. Using transgenic Drosophila expressing human MARK4 (MARK4wt) or a mutant version of MARK4 (MARK4ΔG316E317D), we found that coexpression of MARK4wt and MARK4ΔG316E317D increased total tau levels and enhanced tau-induced neurodegeneration and that MARK4ΔG316E317D had more potent effects than MARK4wt Interestingly, the in vitro kinase activities of MARK4wt and MARK4ΔG316E317D were similar. When tau phosphorylation at Ser-262 and Ser-356 was blocked by alanine substitutions, MARK4wt did not promote tau accumulation or exacerbate neurodegeneration, whereas coexpression of MARK4ΔG316E317D did. Both MARK4wt and MARK4ΔG316E317D increased the levels of oligomeric forms of tau; however, only MARK4ΔG316E317D further increased the detergent insolubility of tau in vivo Together, these findings suggest that MARK4ΔG316E317D increases tau levels and exacerbates tau toxicity via a novel gain-of-function mechanism and that modification in this region of MARK4 may affect disease pathogenesis.
Keywords: Alzheimer's disease (AD); Drosophila; microtubule affinity–regulating kinase 4 (MARK4); neurodegeneration; phosphorylation; tau protein (tau).
© 2020 Oba et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Grundke-Iqbal, I., Iqbal, K., Tung, Y. C., Quinlan, M., Wisniewski, H. M., and Binder, L. I. (1986) Abnormal phosphorylation of the microtubule-associated protein tau (tau) in Alzheimer cytoskeletal pathology. Proc. Natl. Acad. Sci. U. S. A. 83, 4913–4917 10.1073/pnas.83.13.4913 - DOI - PMC - PubMed
-
- Hasegawa, M., Morishima-Kawashima, M., Takio, K., Suzuki, M., Titani, K., and Ihara, Y. (1992) Protein sequence and mass spectrometric analyses of tau in the Alzheimer's disease brain. J. Biol. Chem. 267, 17047–17054 - PubMed
-
- Hanger, D. P., Betts, J. C., Loviny, T. L., Blackstock, W. P., and Anderton, B. H. (1998) New phosphorylation sites identified in hyperphosphorylated tau (paired helical filament-tau) from Alzheimer's disease brain using nanoelectrospray mass spectrometry. J. Neurochem. 71, 2465–2476 10.1046/j.1471-4159.1998.71062465.x - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
