

Altered Outer Hair Cell Mitochondrial and Subsurface Cisternae Connectomics Are Candidate Mechanisms for Hearing Loss in Mice
- PMID: 33020216
- PMCID: PMC7605424
- DOI: 10.1523/JNEUROSCI.2901-19.2020
Altered Outer Hair Cell Mitochondrial and Subsurface Cisternae Connectomics Are Candidate Mechanisms for Hearing Loss in Mice
Abstract
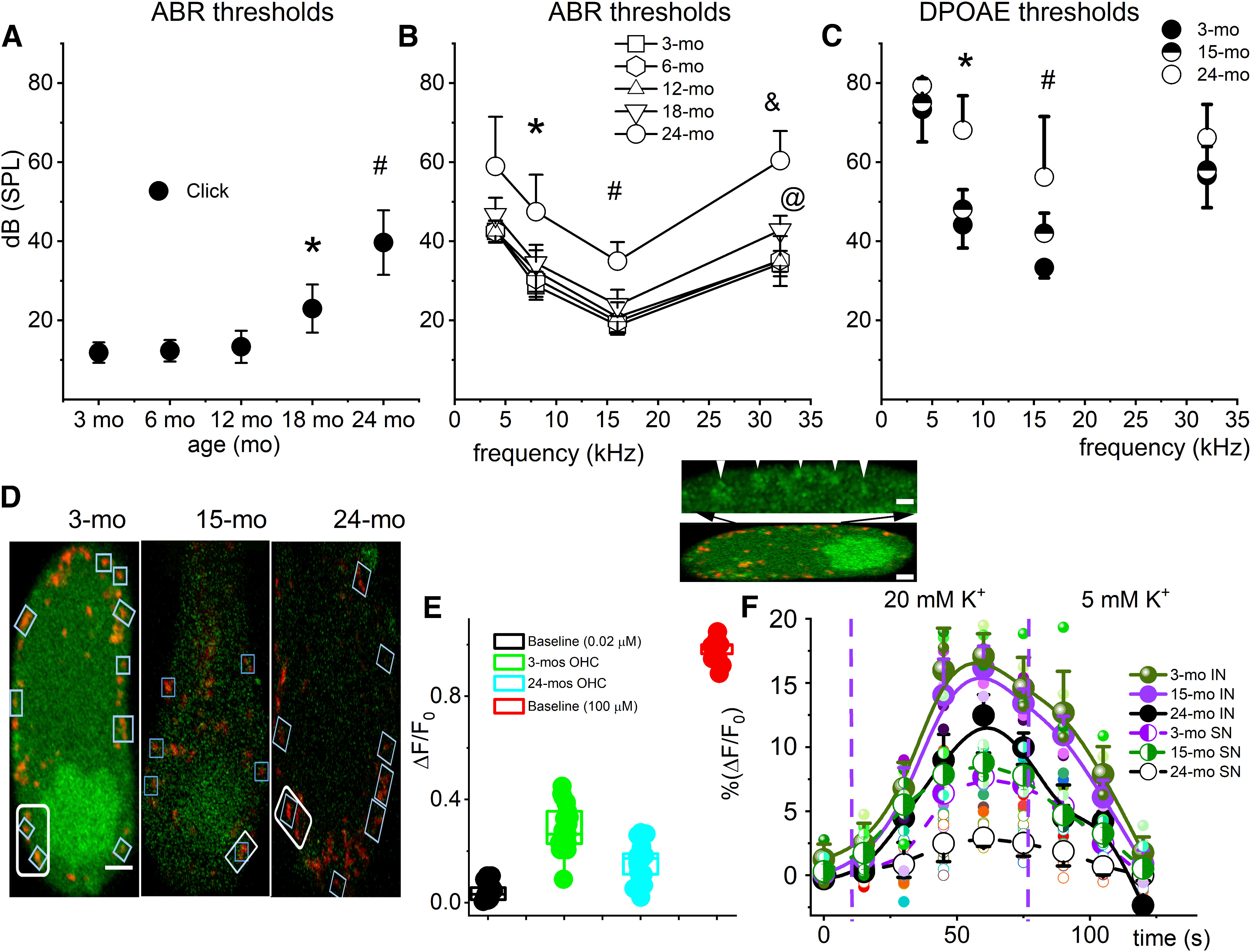
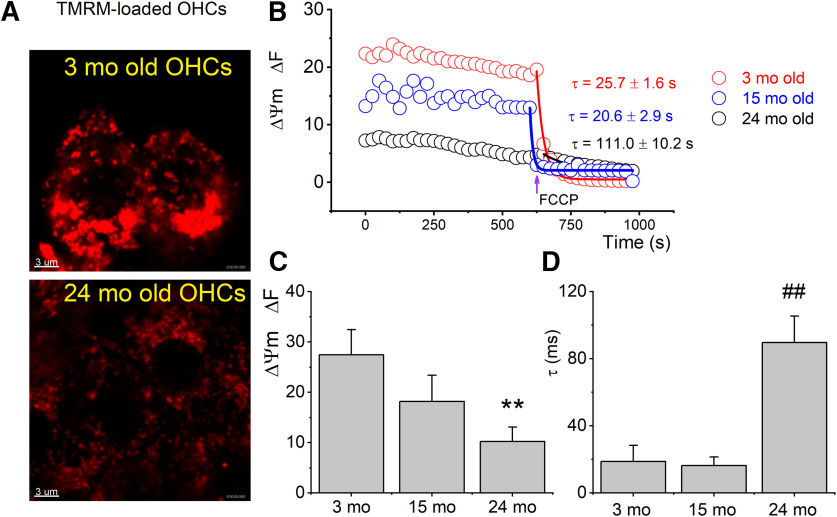
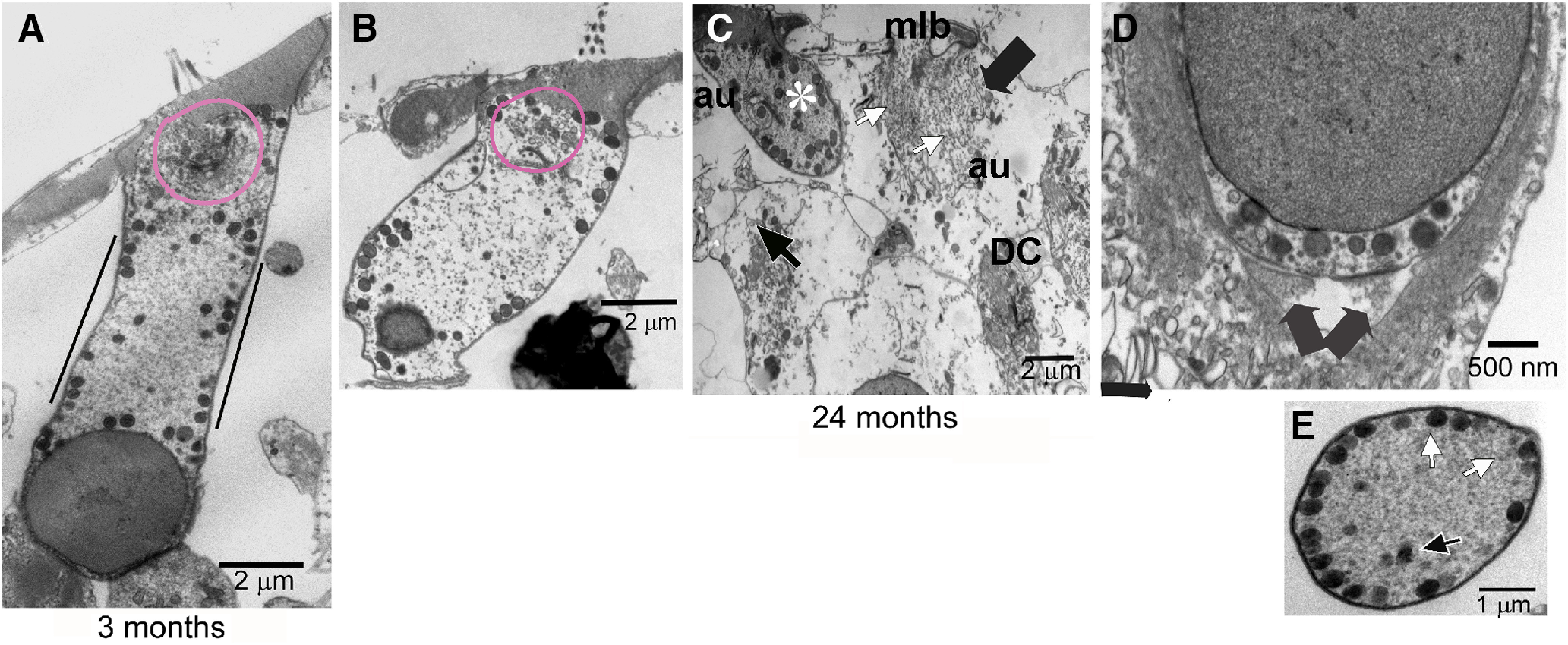
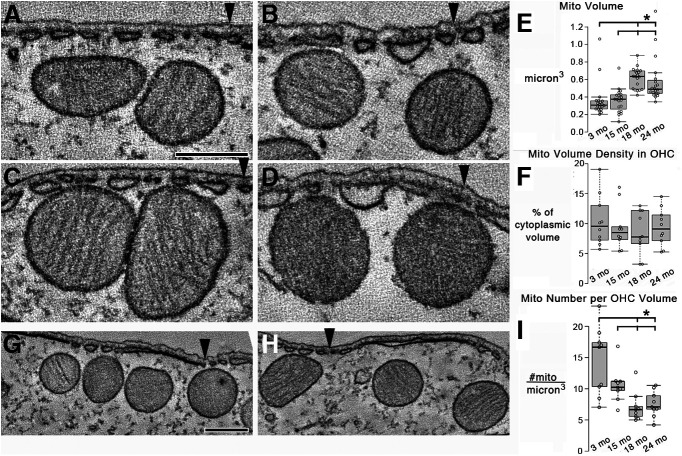
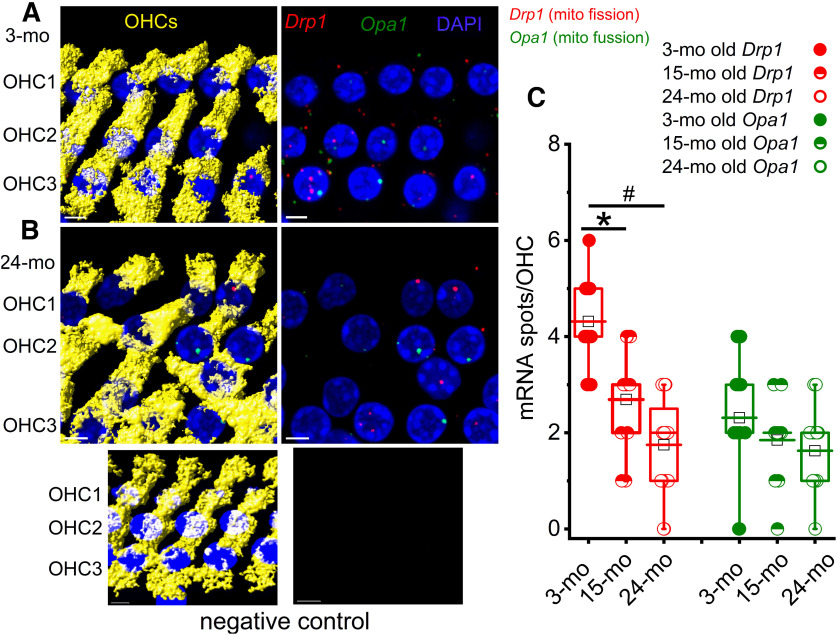
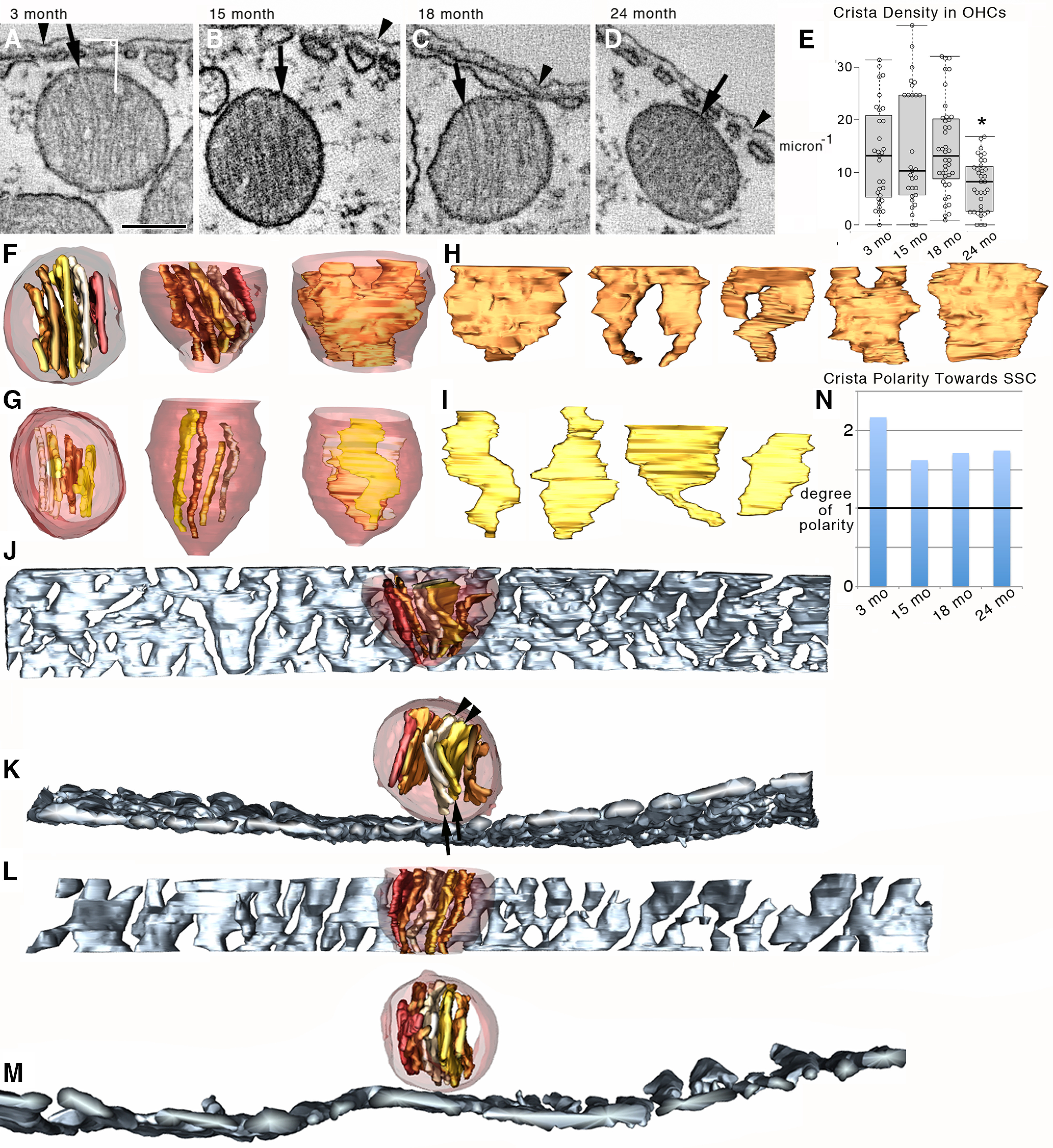
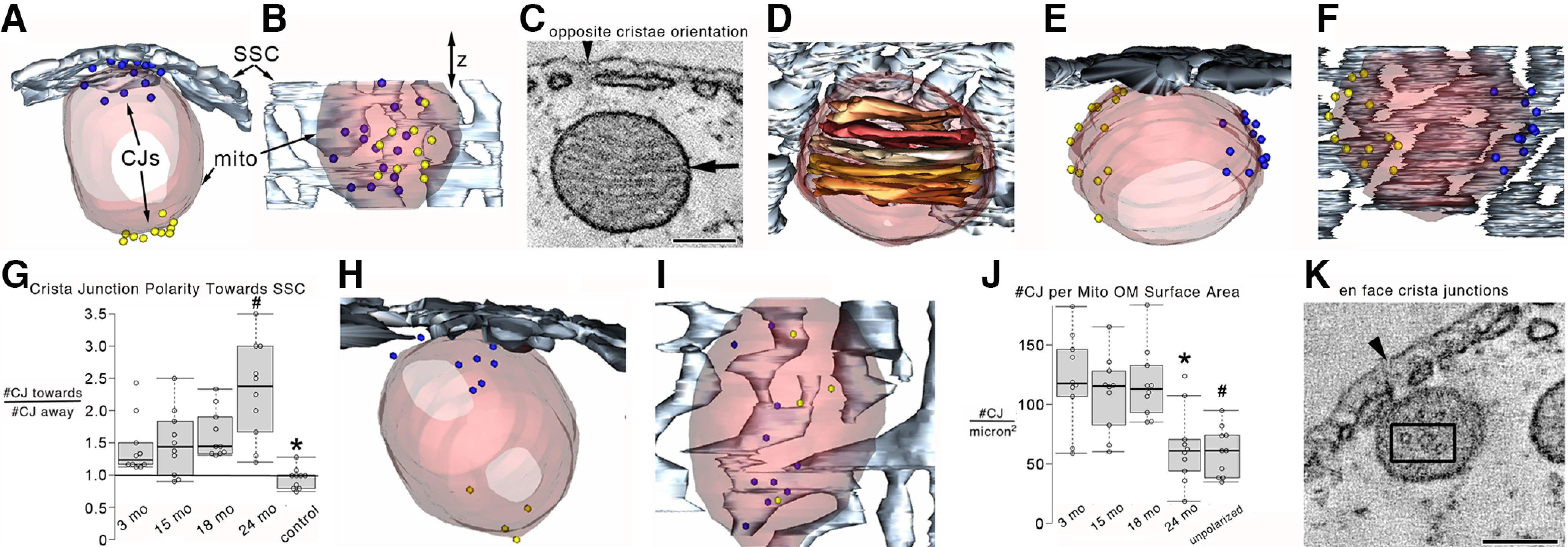
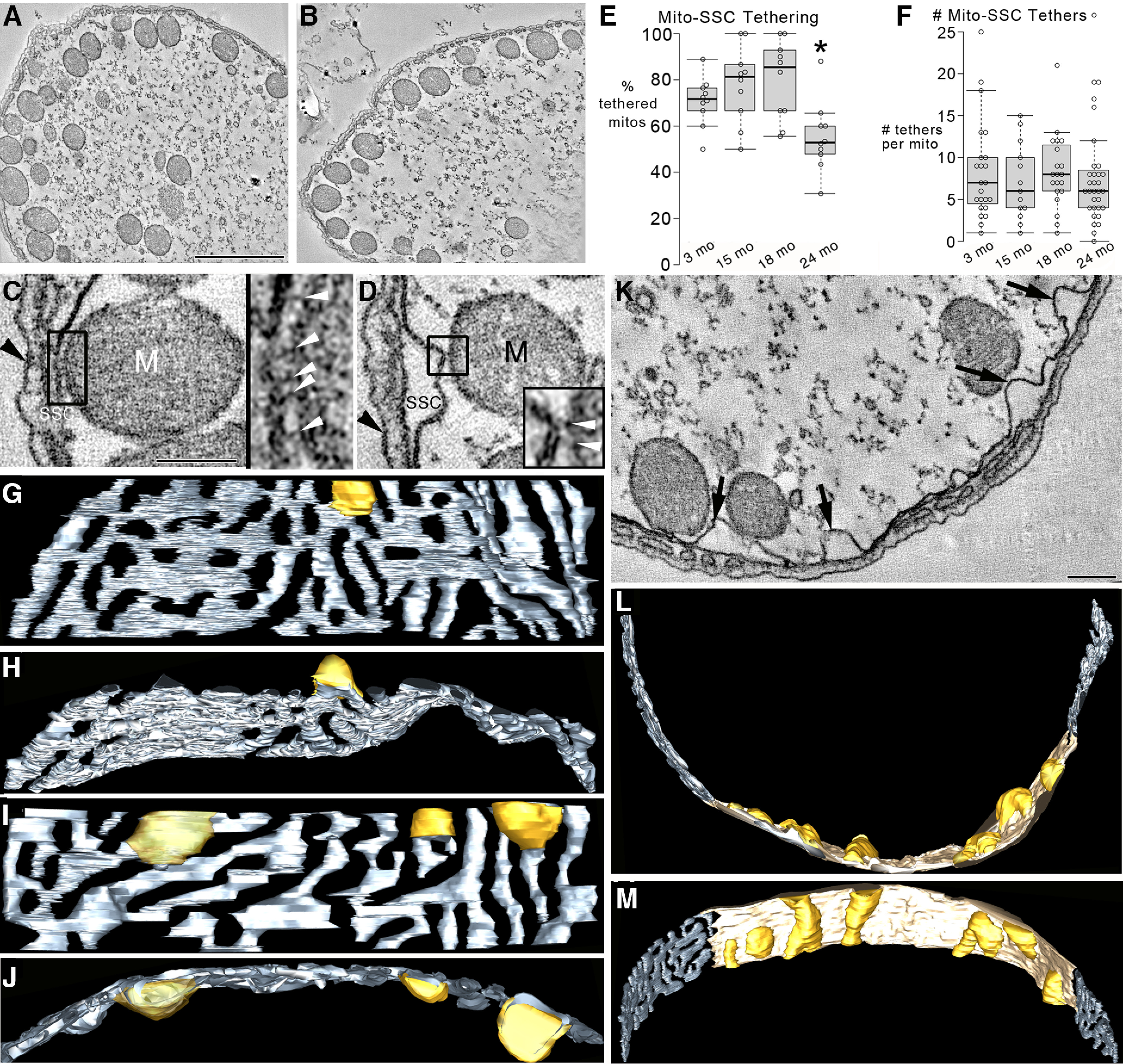
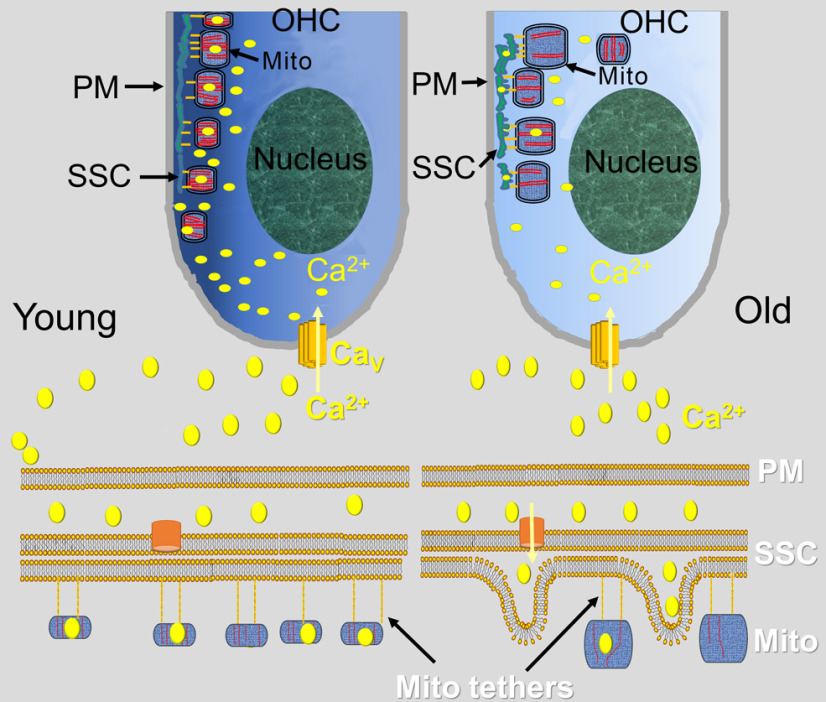
Organelle crosstalk is vital for cellular functions. The propinquity of mitochondria, ER, and plasma membrane promote regulation of multiple functions, which include intracellular Ca2+ flux, and cellular biogenesis. Although the purposes of apposing mitochondria and ER have been described, an understanding of altered organelle connectomics related to disease states is emerging. Since inner ear outer hair cell (OHC) degeneration is a common trait of age-related hearing loss, the objective of this study was to investigate whether the structural and functional coupling of mitochondria with subsurface cisternae (SSC) was affected by aging. We applied functional and structural probes to equal numbers of male and female mice with a hearing phenotype akin to human aging. We discovered the polarization of cristae and crista junctions in mitochondria tethered to the SSC in OHCs. Aging was associated with SSC stress and decoupling of mitochondria with the SSC, mitochondrial fission/fusion imbalance, a remarkable reduction in mitochondrial and cytoplasmic Ca2+ levels, reduced K+-induced Ca2+ uptake, and marked plasticity of cristae membranes. A model of structure-based ATP production predicts profound energy stress in older OHCs. This report provides data suggesting that altered membrane organelle connectomics may result in progressive hearing loss.
Keywords: calcium homeostasis; deafness; hearing loss; inner ear; mitochondria; outer hair cells.
Copyright © 2020 the authors.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Early functional alterations in membrane properties and neuronal degeneration are hallmarks of progressive hearing loss in NOD mice.Sci Rep. 2019 Aug 20;9(1):12128. doi: 10.1038/s41598-019-48376-x. Sci Rep. 2019. PMID: 31431657 Free PMC article.
-
Chronic Electro-Acoustic Stimulation May Interfere With Electric Threshold Recovery After Cochlear Implantation in the Aged Guinea Pig.Ear Hear. 2024 Nov-Dec 01;45(6):1554-1567. doi: 10.1097/AUD.0000000000001545. Epub 2024 Jul 12. Ear Hear. 2024. PMID: 38992863 Free PMC article.
-
The local translation of KNa in dendritic projections of auditory neurons and the roles of KNa in the transition from hidden to overt hearing loss.Aging (Albany NY). 2019 Dec 8;11(23):11541-11564. doi: 10.18632/aging.102553. Epub 2019 Dec 8. Aging (Albany NY). 2019. PMID: 31812952 Free PMC article.
-
Ventilation tubes (grommets) for otitis media with effusion (OME) in children.Cochrane Database Syst Rev. 2023 Nov 15;11(11):CD015215. doi: 10.1002/14651858.CD015215.pub2. Cochrane Database Syst Rev. 2023. PMID: 37965944 Free PMC article.
-
Bilateral versus unilateral hearing aids for bilateral hearing impairment in adults.Cochrane Database Syst Rev. 2017 Dec 19;12(12):CD012665. doi: 10.1002/14651858.CD012665.pub2. Cochrane Database Syst Rev. 2017. PMID: 29256573 Free PMC article.
Cited by
-
Mitochondrial morphology governs ATP production rate.J Gen Physiol. 2023 Sep 4;155(9):e202213263. doi: 10.1085/jgp.202213263. Epub 2023 Aug 24. J Gen Physiol. 2023. PMID: 37615622 Free PMC article.
-
A Cristae-Like Microcompartment in Desulfobacterota.mBio. 2022 Dec 20;13(6):e0161322. doi: 10.1128/mbio.01613-22. Epub 2022 Nov 2. mBio. 2022. PMID: 36321837 Free PMC article.
-
Morphological principles of neuronal mitochondria.J Comp Neurol. 2022 Apr;530(6):886-902. doi: 10.1002/cne.25254. Epub 2021 Nov 1. J Comp Neurol. 2022. PMID: 34608995 Free PMC article.
-
Presbycusis: Pathology, Signal Pathways, and Therapeutic Strategy.Adv Sci (Weinh). 2025 Aug;12(29):e2410413. doi: 10.1002/advs.202410413. Epub 2025 May 11. Adv Sci (Weinh). 2025. PMID: 40349177 Free PMC article. Review.
-
Peripheral Auditory Nerve Impairment in a Mouse Model of Syndromic Autism.J Neurosci. 2022 Oct 19;42(42):8002-8018. doi: 10.1523/JNEUROSCI.0253-22.2022. Epub 2022 Sep 30. J Neurosci. 2022. PMID: 36180228 Free PMC article.
References
-
- Babu V, Vazirian S, Perkins G, Lysakowski A (2017) Energy output of mitochondria located near synaptic ribbons in inner ear hair cells. FASEB J 31:740–21..
-
- Bai U, Seidman MD, Hinojosa R, Quirk WS (1997) Mitochondrial DNA deletions associated with aging and possibly presbycusis: a human archival temporal bone study. Am J Otol 18:449–453. - PubMed
-
- Bawa B, Abbott LC (2008) Analysis of calcium ion homeostasis and mitochondrial function in cerebellar granule cells of adult CaV 2.1 calcium ion channel mutant mice. Neurotox Res 13:1–18. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous