

Extracellular matrix plasticity as a driver of cell spreading
- PMID: 33020289
- PMCID: PMC7584996
- DOI: 10.1073/pnas.2008801117
Extracellular matrix plasticity as a driver of cell spreading
Abstract
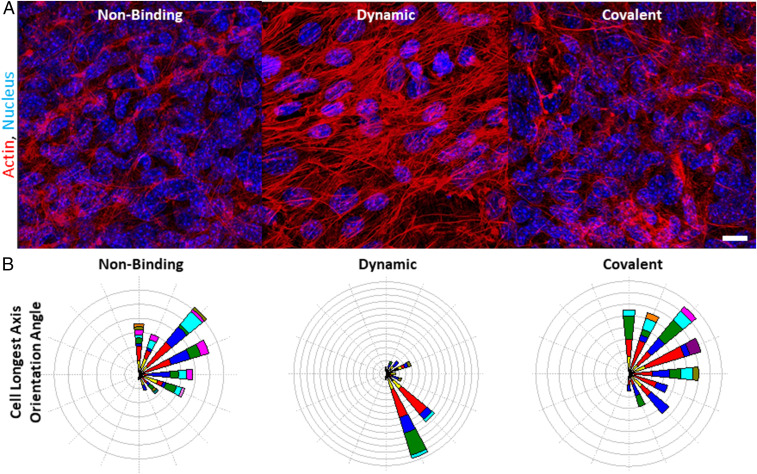
Mammalian cell morphology has been linked to the viscoelastic properties of the adhesion substrate, which is particularly relevant in biological processes such as wound repair and embryonic development where cell spreading and migration are critical. Plastic deformation, degradation, and relaxation of stress are typically coupled in biomaterial systems used to explore these effects, making it unclear which variable drives cell behavior. Here we present a nondegradable polymer architecture that specifically decouples irreversible creep from stress relaxation and modulus. We demonstrate that network plasticity independently controls mesenchymal stem cell spreading through a biphasic relationship dependent on cell-intrinsic forces, and this relationship can be shifted by inhibiting actomyosin contractility. Kinetic Monte Carlo simulations also show strong correlation with experimental cell spreading data as a function of the extracellular matrix (ECM) plasticity. Furthermore, plasticity regulates many ECM adhesion and remodeling genes. Altogether, these findings confirm a key role for matrix plasticity in stem cell biophysics, and we anticipate this will have ramifications in the design of biomaterials to enhance therapeutic applications of stem cells.
Keywords: biomaterials; extracellular matrix; plasticity; stem cell; viscoelasticity.
Conflict of interest statement
The authors declare no competing interest.
Figures





References
-
- Li Q. et al. ., Extracellular matrix scaffolding guides lumen elongation by inducing anisotropic intercellular mechanical tension. Nat. Cell Biol. 18, 311–318 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
