

One ring to bring them all and in the darkness bind them: The trafficking of heme without deliverers
- PMID: 33022276
- PMCID: PMC7756907
- DOI: 10.1016/j.bbamcr.2020.118881
One ring to bring them all and in the darkness bind them: The trafficking of heme without deliverers
Abstract
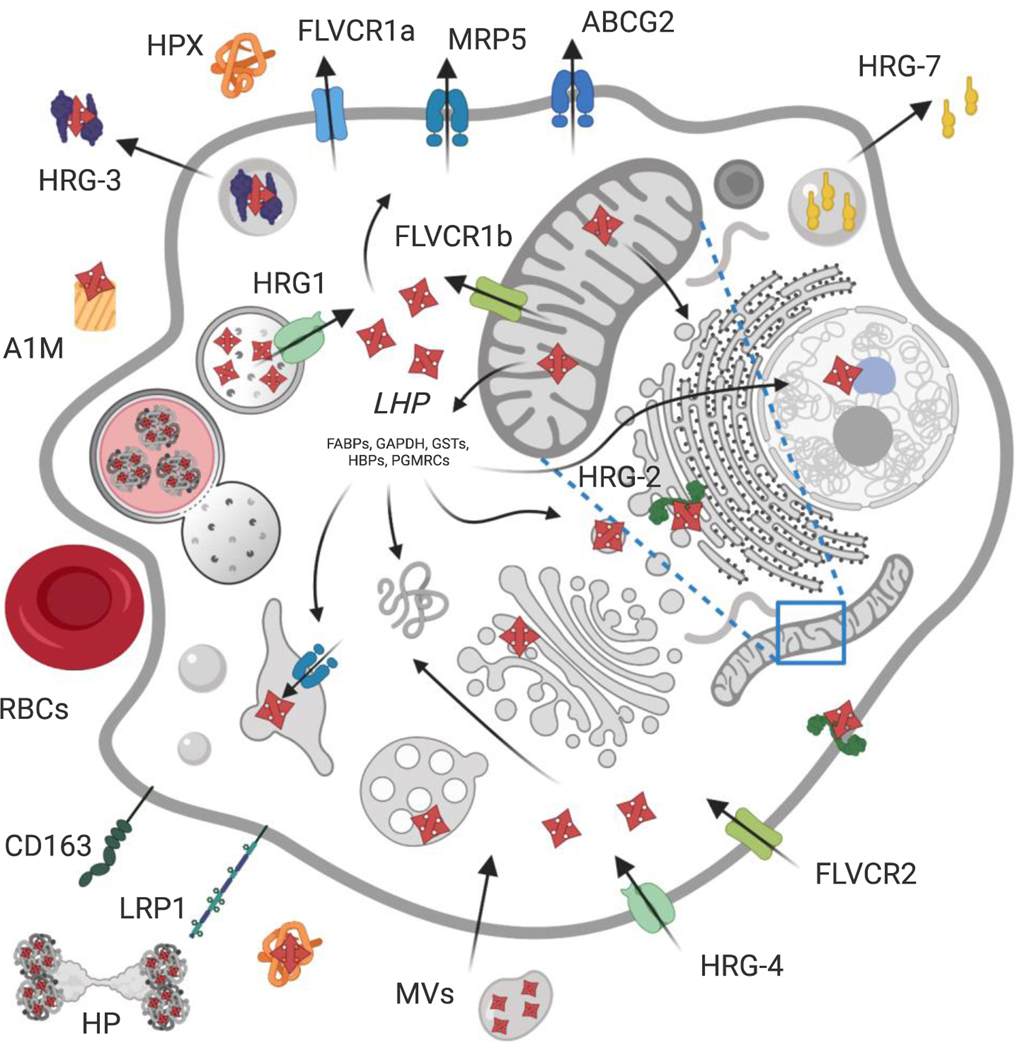
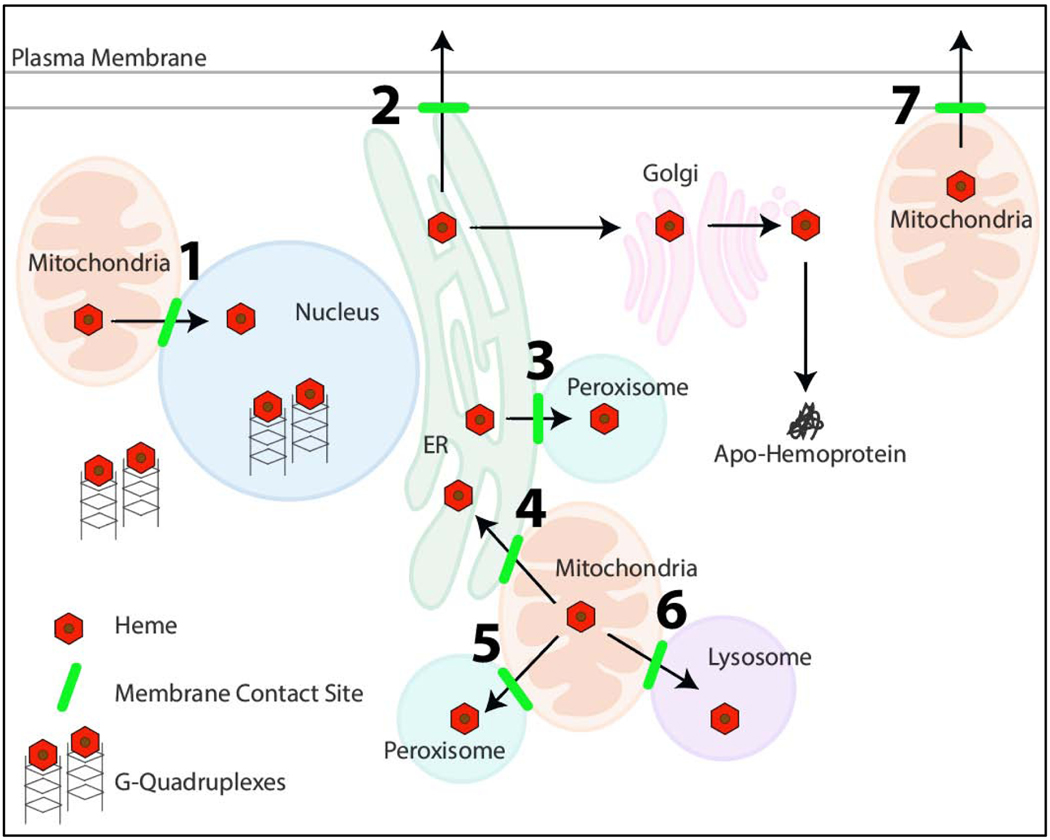
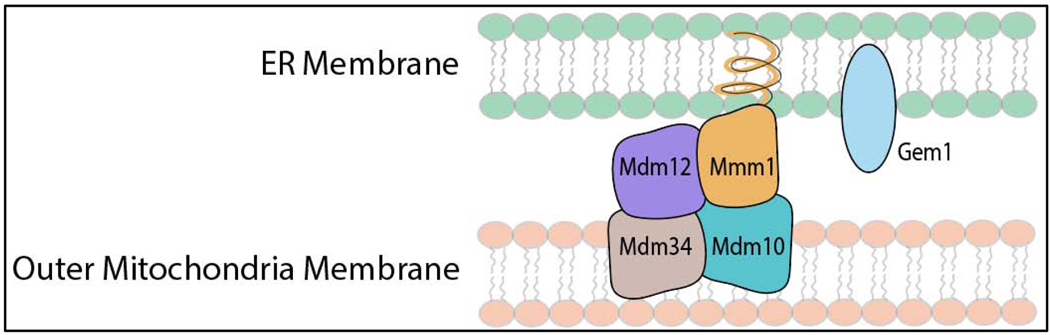
Heme, as a hydrophobic iron-containing organic ring, is lipid soluble and can interact with biological membranes. The very same properties of heme that nature exploits to support life also renders heme potentially cytotoxic. In order to utilize heme, while also mitigating its toxicity, cells are challenged to tightly control the concentration and bioavailability of heme. On the bright side, it is reasonable to envision that, analogous to other transition metals, a combination of membrane-bound transporters, soluble carriers, and chaperones coordinate heme trafficking to subcellular compartments. However, given the dual properties exhibited by heme as a transition metal and lipid, it is compelling to consider the dark side: the potential role of non-proteinaceous biomolecules including lipids and nucleic acids that bind, sequester, and control heme trafficking and bioavailability. The emergence of inter-organellar membrane contact sites, as well as intracellular vesicles derived from various organelles, have raised the prospect that heme can be trafficked through hydrophobic channels. In this review, we aim to focus on heme delivery without deliverers - an alternate paradigm for the regulation of heme homeostasis through chaperone-less pathways for heme trafficking.
Keywords: Heme; Iron; Porphyrin; Tetrapyrrole; Trafficking.
Copyright © 2020 Elsevier B.V. All rights reserved.
Figures
References
-
- Sassa S, Why heme needs to be degraded to iron, biliverdin IXalpha, and carbon monoxide?, Antioxid Redox Signal, 6 (2004) 819–824. - PubMed
-
- Reedy CJ, Gibney BR, Heme protein assemblies, Chem Rev, 104 (2004) 617–649. - PubMed
-
- Kumar S, Bandyopadhyay U, Free heme toxicity and its detoxification systems in human, Toxicol Lett, 157 (2005) 175–188. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
