

Gαs directly drives PDZ-RhoGEF signaling to Cdc42
- PMID: 33023908
- PMCID: PMC7863908
- DOI: 10.1074/jbc.AC120.015204
Gαs directly drives PDZ-RhoGEF signaling to Cdc42
Abstract
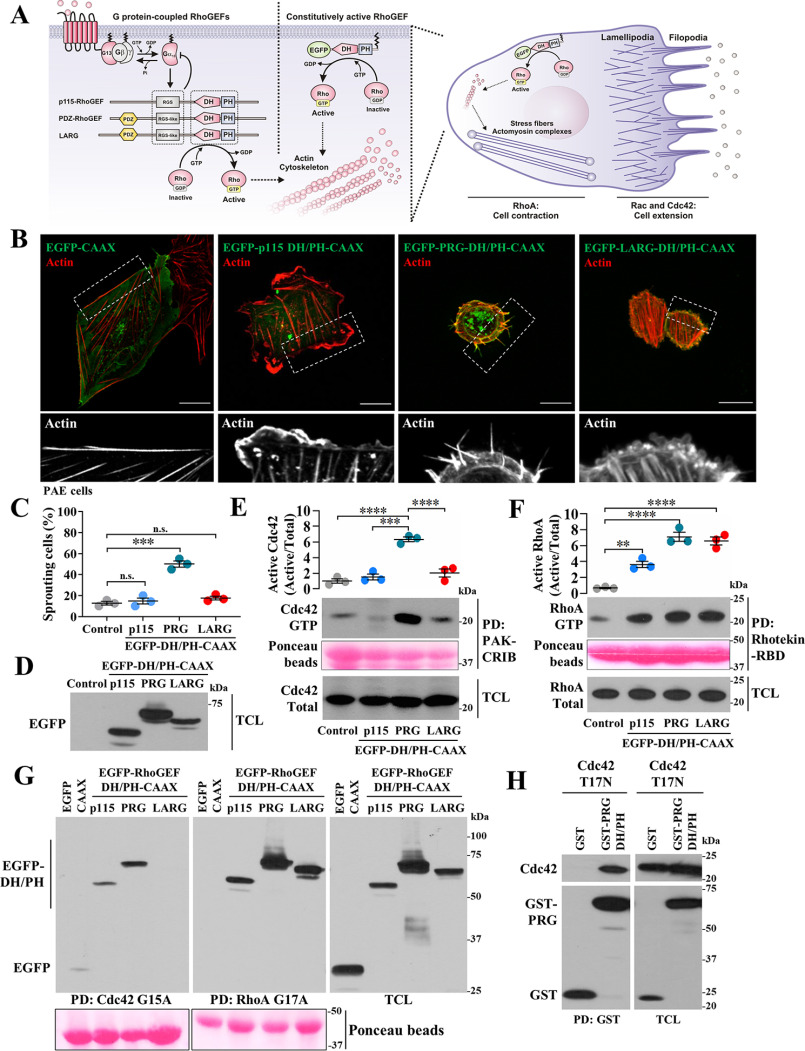
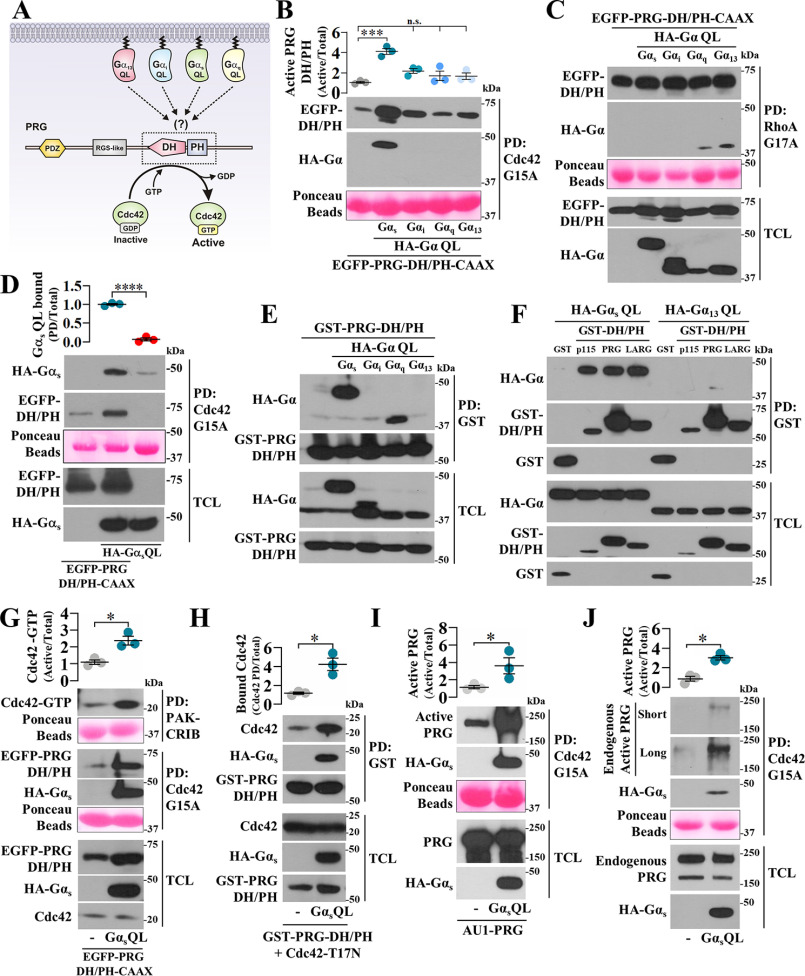
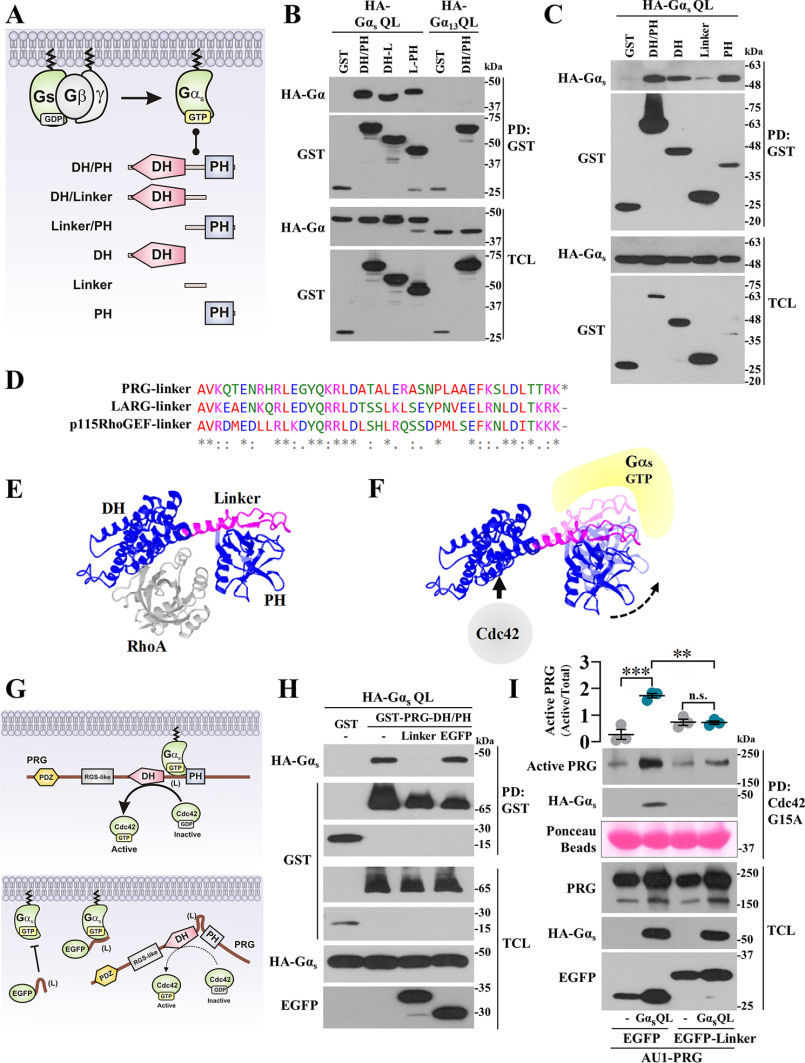
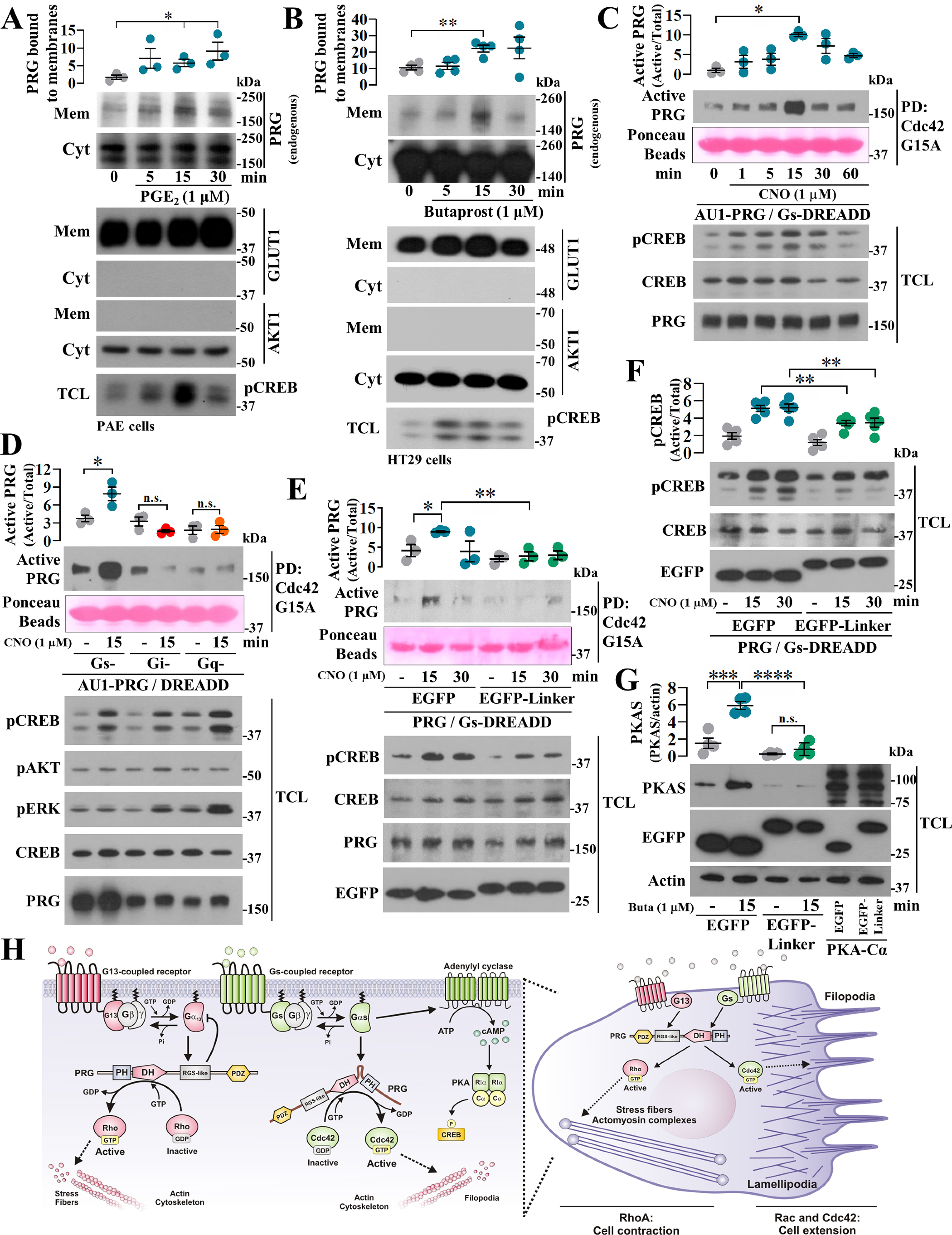
Gα proteins promote dynamic adjustments of cell shape directed by actin-cytoskeleton reorganization via their respective RhoGEF effectors. For example, Gα13 binding to the RGS-homology (RH) domains of several RH-RhoGEFs allosterically activates these proteins, causing them to expose their catalytic Dbl-homology (DH)/pleckstrin-homology (PH) regions, which triggers downstream signals. However, whether additional Gα proteins might directly regulate the RH-RhoGEFs was not known. To explore this question, we first examined the morphological effects of expressing shortened RH-RhoGEF DH/PH constructs of p115RhoGEF/ARHGEF1, PDZ-RhoGEF (PRG)/ARHGEF11, and LARG/ARHGEF12. As expected, the three constructs promoted cell contraction and activated RhoA, known to be downstream of Gα13 Intriguingly, PRG DH/PH also induced filopodia-like cell protrusions and activated Cdc42. This pathway was stimulated by constitutively active Gαs (GαsQ227L), which enabled endogenous PRG to gain affinity for Cdc42. A chemogenetic approach revealed that signaling by Gs-coupled receptors, but not by those coupled to Gi or Gq, enabled PRG to bind Cdc42. This receptor-dependent effect, as well as CREB phosphorylation, was blocked by a construct derived from the PRG:Gαs-binding region, PRG-linker. Active Gαs interacted with isolated PRG DH and PH domains and their linker. In addition, this construct interfered with GαsQ227L's ability to guide PRG's interaction with Cdc42. Endogenous Gs-coupled prostaglandin receptors stimulated PRG binding to membrane fractions and activated signaling to PKA, and this canonical endogenous pathway was attenuated by PRG-linker. Altogether, our results demonstrate that active Gαs can recognize PRG as a novel effector directing its DH/PH catalytic module to gain affinity for Cdc42.
Keywords: ARHGEF11; Cdc42; DH/PH catalytic module; G protein–coupled receptor (GPCR); GPCR; Galpha-s; Gαs; PDZ-RhoGEF; PDZ-RhoGEF (PRG); Rho (Rho GTPase); Rho GTPases; Rho guanine nucleotide exchange factor (RhoGEF); cell signaling; guanine nucleotide exchange factor (GEF); heterotrimeric G protein.
© 2020 Castillo-Kauil et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
Comment in
-
A Gs-RhoGEF interaction: An old G protein finds a new job.J Biol Chem. 2020 Dec 11;295(50):16929-16930. doi: 10.1074/jbc.H120.016606. J Biol Chem. 2020. PMID: 33310745 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
