

Review
doi: 10.1038/s41577-020-00446-2.
Epub 2020 Oct 6.
Transcriptional regulation of memory B cell differentiation
Affiliations
- PMID: 33024284
- PMCID: PMC7538181
- DOI: 10.1038/s41577-020-00446-2
Item in Clipboard
Review
Transcriptional regulation of memory B cell differentiation
Nat Rev Immunol.
2021 Apr.
Abstract
Memory B cells (MBCs) are critical for the rapid development of protective immunity following re-infection. MBCs capable of neutralizing distinct subclasses of pathogens, such as influenza and HIV, have been identified in humans. However, efforts to develop vaccines that induce broadly protective MBCs to rapidly mutating pathogens have not yet been successful. Better understanding of the signals regulating MBC development and function are essential to overcome current challenges hindering successful vaccine development. Here, we discuss recent advancements regarding the signals and transcription factors regulating germinal centre-derived MBC development and function.
Conflict of interest statement
The authors declare no competing interests.
Figures
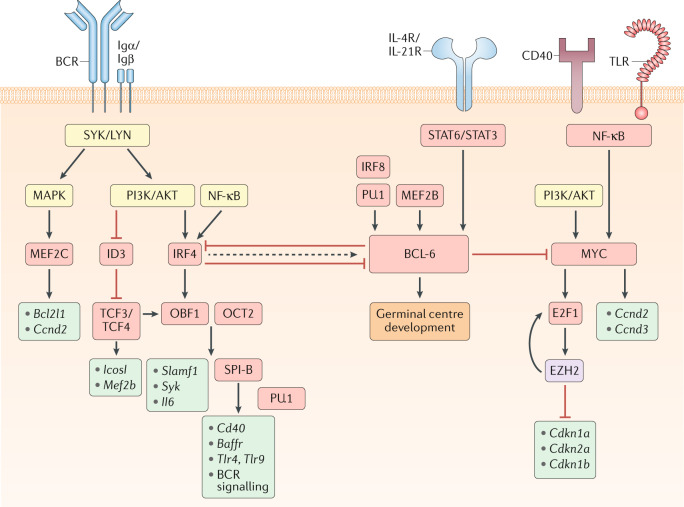
Model for signalling pathways and transcription factors that regulate B cell commitment to the germinal centre (GC) fate. Boxes that indicate signalling molecules are coloured yellow, transcription regulators red, downstream gene targets turquoise and epigenetic modifiers purple. The B cell receptor (BCR), via its signalling subunits Igα and Igβ as well as downstream tyrosine kinases such as SYK and LYN, activates the mitogen-activated protein kinase (MAPK) and phosphoinositide 3-kinase (PI3K)/AKT pathways. MAPK signalling induces the expression of the transcriptional activator myocyte enhancer binding factor 2c (MEF2C), which promotes the transcription of B cell lymphoma-extra large (Bcl2l1) and cyclin D2 (Ccnd2). The expression of the transcriptional repressor inhibitor of DNA binding 3 (ID3) is reduced following B cell activation, allowing for transcription factor 3/4 (TCF3/4)-driven induction of Icosl and Mef2b transcription. PI3K/AKT and nuclear factor-κB (NF-κB) signalling also induce the expression of the transcription factor interferon regulatory factor 4 (IRF4). IRF4 and TCF3/4 induce the expression of the co-activator OBF1 (also known as POU class 2 homeobox associating factor 1 (POU2AF1)), which cooperates with OCT2 (also known as POU class 2 homeobox 2 (POU2F2)) to promote transcription of Slamf1, Syk and Il6. OBF1 and OCT2 can also induce the expression of the transcription factor SPI-B, which acts in a redundant fashion with PU.1 to enhance the transcription of genes encoding B cell surface receptors, such as Cd40, B cell activating factor receptor (Baffr), Toll-like receptor 4 (Tlr4) and Tlr9, and many components of the BCR signalling pathway, including Blnk and Btk. Although transiently elevated levels of IRF4 can induce the expression of B cell lymphoma 6 (BCL-6), sustained IRF4 levels will repress BCL-6 expression. BCL-6 expression is also induced by the transcription factors MEF2B and IRF8/PU.1 as well as the cytokines IL-4 and IL-21, which bind to their respective receptors (IL-4R and IL-21R) and induce signal transducer and activator of transcription 6 (STAT6)/STAT3 signalling. CD40 and/or TLR-driven NF-κB signalling, alongside PI3K/AKT signalling, will induce the expression of the transcription factor MYC, which promotes cellular proliferation by inducing the transcription of Ccnd2/Ccnd3 and the expression of the transcription factor E2F transcription factor 1 (E2F1). E2F1 induces expression of Ezh2 that encodes a Polycomb repressive complex 2 (PRC2) enzymatic component. Enhancer of zeste homologue 2 (EZH2) promotes cell cycle progression by repressing the expression of Cdkn1a, Cdkn2a and Cdkn1b, which encode cyclin-dependent kinase inhibitors. EZH2 also promotes E2F1 release from the retinoblastoma (Rb) protein via phosphorylation of Rb, thereby enhancing E2F1 activation and further EZH2 expression. BCL-6 can directly repress MYC expression, thereby limiting the number of cell divisions that GC B cells undergo. BCL-6 promotes GC B cell development through regulation of numerous genes controlling cellular processes including the DNA damage response, B cell migration, apoptosis, BCR and CD40 signalling, plasma cell differentiation and T cell:B cell interactions. Together, these transcriptional regulators allow for the precise control of GC initiation that is necessary to balance the competing needs of the immune system to induce a protective response while limiting immunopathology.
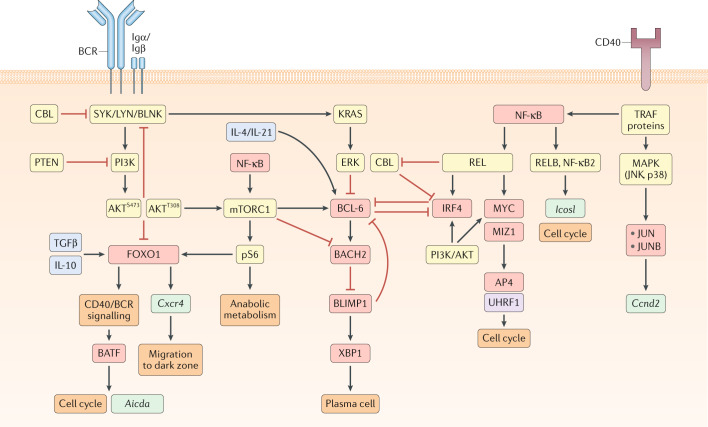
Model for signalling pathways and transcription factors that regulate germinal centre (GC) B cell maintenance. Boxes that indicate signalling molecules are coloured yellow, transcription factors red, downstream gene targets turquoise, cytokines blue (drawn inside cell for simplicity) and epigenetic modifiers purple. The B cell receptor (BCR), via its signalling subunits Igα and Igβ, engages the tyrosine kinases SYK and LYN and the adapter protein BLNK, leading to activation of phosphoinositide 3-kinase (PI3K) and subsequent phosphorylation of AKT at S473. In GC B cells, AKT is already phosphorylated at T308. PI3K/AKT signalling represses forkhead box O1 (FOXO1) expression and attenuates BCR signalling through phosphorylation of the signalling molecules SHP1, CSK and HPK1, which negatively regulate the activity of SYK, LYN and BLNK, respectively. GC B cells also express high levels of the phosphatase and tensin homologue (PTEN), which restrains PI3K-mediated phosphorylation of AKT at S473. PI3K/AKT signalling, along with CD40-driven nuclear factor-κB (NF-κB) activity, can induce mechanistic target of rapamycin complex 1 (mTORC1) signalling and lead to the phosphorylation of the ribosomal protein S6 (pS6) and the adoption of an anabolic metabolic state through the increased accumulation of biomass. mTORC1 can also promote the expression of B cell lymphoma 6 (BCL-6) and repress BTB domain and CNC homology 2 (BACH2) expression. pS6, alongside IL-10 and transforming growth factor-β (TGFβ), can induce the expression of the transcription factor FOXO1. FOXO1 promotes the transcription of Cxcr4, which facilitates GC B cell migration to the dark zone. FOXO1 also promotes CD40/BCR signalling, indirectly resulting in expression of the transcription factor basic leucine zipper transcription factor (BATF). BATF directly promotes the transcription of Aicda and cell cycle progression, likely through interaction with other transcription factors of the JUN or interferon regulatory factor (IRF) families. BCR signalling also engages the GTPase KRAS, which activates the kinase ERK, resulting in repressed BCL-6 expression. CD40 signalling leads to the recruitment of the kinase MEKK1 to the adapter molecule TRAF2 and results in activation of the mitogen-activated protein kinases (MAPKs) JNK and p38. MAPK signalling induces the expression of the transcription factors JUN and JUNB, as well as the transcription of Ccnd2. CD40 signalling also signals through TRAF proteins to induce NF-κB. NF-κB signals through both the canonical (REL) and non-canonical (RELB, NF-κB2) pathways to regulate GC maintenance. RELB and NF-κB2 induce Icosl expression and promote the induction of the cell cycle. The precise mechanisms by which RELB and NF-κB2 regulate the cell cycle is unclear but may relate to induction of ICOSL expression. REL and PI3K/AKT signalling induce the expression of IRF4 and MYC. MYC acts in a complex with MYC-interacting zinc finger protein 1 (MIZ1) to induce the expression of activating enhancer binding protein 4 (AP4) and the epigenetic regulator ubiquitin-like, containing PHD and RING finger domains, 1 (UHRF1), both of which promote the cell cycle. Strong canonical NF-κB signalling also represses expression of the ubiquitin ligase CBL (not shown). CBL inhibits plasma cell differentiation through repression of IRF4 expression (not shown) and BCR signalling via degradation of SYK. IRF4 and BCL-6 are mutually antagonistic transcription factors. BCL-6 induces BACH2 expression, which represses B lymphocyte-induced maturation protein 1 (BLIMP1) expression. In turn, BLIMP1 can repress BCL-6 expression and induce XBP1 expression, resulting in plasma cell differentiation.
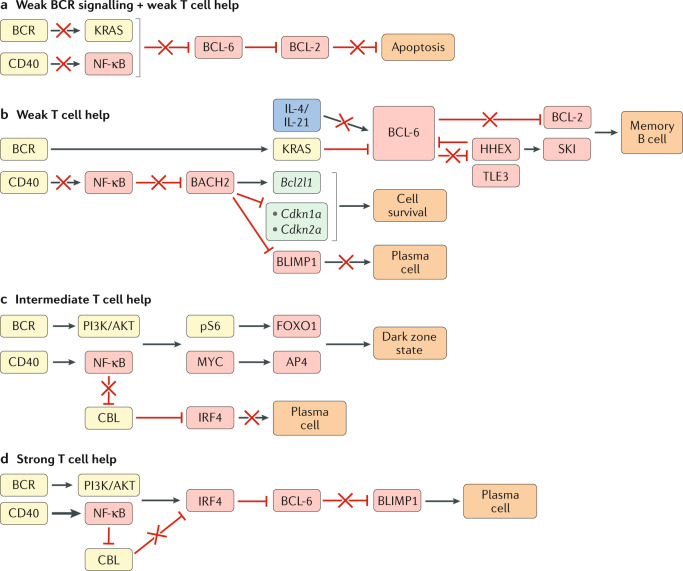
Germinal centre (GC) B cell differentiation is regulated by the extent of signals received by the B cell through the antigen-engaged B cell receptor (BCR) and CD40 (via CD40L-expressing T cells). Boxes indicating signalling molecules are coloured yellow, transcription factors red, cytokines blue and downstream gene targets turquoise. A red cross indicates a pathway that is not functional under the condition illustrated. a | Weak BCR signalling and weak T cell help results in a failure of GC B cells to receive sufficient signals through the BCR and CD40 to induce KRAS and nuclear factor-κB (NF-κB)-driven repression of B cell lymphoma 6 (BCL-6) expression. Repression of BCL-2 expression by BCL-6 in these cells will result in apoptosis owing to a failure to receive sufficient survival signals. b | GC B cells that receive weak T cell help (transmitted via CD40) will not receive sufficient CD40-induced NF-κB signals to indirectly repress BTB domain and CNC homology 2 (BACH2). BACH2 promotes GC B cell survival through induction of the anti-apoptotic gene Bcl2l1 and repression of cyclin-dependent kinase inhibitor family genes (Cdkn1a, Cdkn2a). BACH2 also represses B lymphocyte-induced maturation protein 1 (BLIMP1) and, accordingly, restricts plasma cell differentiation. The continued survival of B cells that receive weak T cell help is dependent on receiving sufficient BCR-driven KRAS signalling to repress BCL-6 expression. Exposure to T cell-derived IL-4 and IL-21 normally stabilizes BCL-6 expression in GC B cells. However, B cells that receive weak T cell-derived CD40 will also have low exposure to IL-4 and IL-21, resulting in impaired BCL-6 expression and induction of BCL-6-repressed genes such as haematopoietically expressed homeobox (Hhex) and Bcl2. HHEX interacts with the co-repressor transducin-like enhancer 3 (TLE3) to further repress BCL-6 expression and promote memory B cell development through the upregulation of BCL-2 and the transcription factor Sloan-Kettering Institute (SKI). c | GC B cells that receive intermediate T cell help receive both phosphoinositide 3-kinase (PI3K)/AKT and NF-κB signalling, resulting in phosphorylation of the ribosomal protein S6 (pS6) and expression of MYC. pS6 and MYC subsequently induce expression of forkhead box O1 (FOXO1) and activating enhancer binding protein 4 (AP4), respectively, ultimately leading to the adoption of the highly proliferative dark zone state. Intermediate levels of CD40 signalling do not drive sufficient NF-κB signalling to repress expression of the ubiquitin ligase CBL, allowing CBL to continue to repress interferon regulatory factor 4 (IRF4) expression and prevent plasma cell differentiation. d | GC B cells that receive strong T cell help receive both PI3K/AKT and NF-κB signalling. Strong NF-κB signalling represses CBL expression and allows for upregulation of IRF4. IRF4 represses BCL-6 expression and leads to expression of BLIMP1 and plasma cell differentiation.
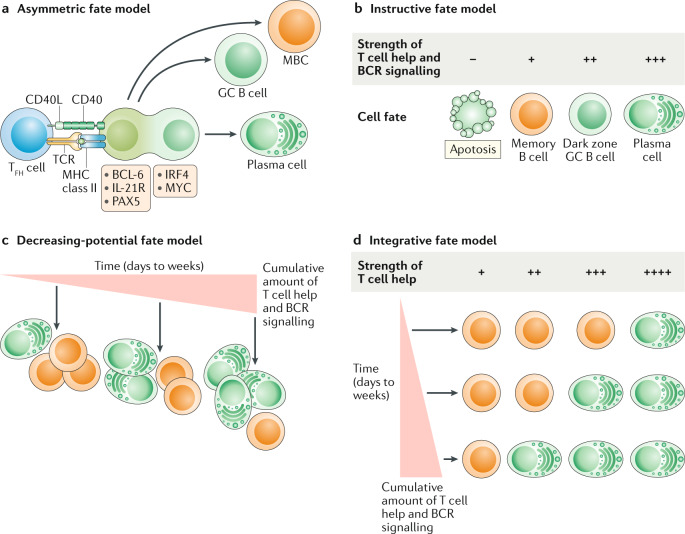
a | Asymmetric fate model. Germinal centre (GC) B cells that interact with follicular helper T (TFH) cells will undergo cellular polarization, resulting in unequal distribution of fate-altering molecules by daughter cells. Daughter cells inheriting fate-altering molecules that promote the expression of interferon regulatory factor 4 (IRF4) and MYC preferentially differentiate into plasma cells, whereas cells inheriting B cell lymphoma 6 (BCL-6), PAX5 and IL-21R either maintain GC B cell identity or differentiate into memory B cells (MBCs). b | Instructive fate model. The strength of T cell help and B cell receptor (BCR) signalling received by GC B cells dictates cell fate. Strong T cell help favours plasma cell differentiation and the dark zone state, whereas weak T cell help predisposes GC B cells to differentiate into MBCs or to undergo apoptosis. c | Decreasing-potential fate model. GC B cell fate is determined by the cumulative strength of T cell help and BCR signalling, accrued over the length of duration in the GC (days to weeks). As GC B cells undergo repeated rounds of cyclic re-entry, they progressively lose their ability to differentiate into MBCs, resulting in a shift towards increased plasma cell differentiation at late GC time points. d | Integrative fate model. GC B cells integrate both the current quality and the cumulative amounts of received signals when making cell fate decisions. GC B cells that have received less cumulative T cell help and BCR signalling need a stronger signal to differentiate into plasma cells. Conversely, GC B cells that have been in the GC longer and received more cumulative signals require a weaker signal to induce plasma cell differentiation. CD40L, CD40 ligand; MHC, major histocompatibility complex; TCR, T cell receptor.
References
-
- Dogan I, et al. Multiple layers of B cell memory with different effector functions. Nat. Immunol. 2009;10:1292–1299. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
