

Acidobacteria are active and abundant members of diverse atmospheric H2-oxidizing communities detected in temperate soils
- PMID: 33024291
- PMCID: PMC8027828
- DOI: 10.1038/s41396-020-00750-8
Acidobacteria are active and abundant members of diverse atmospheric H2-oxidizing communities detected in temperate soils
Abstract
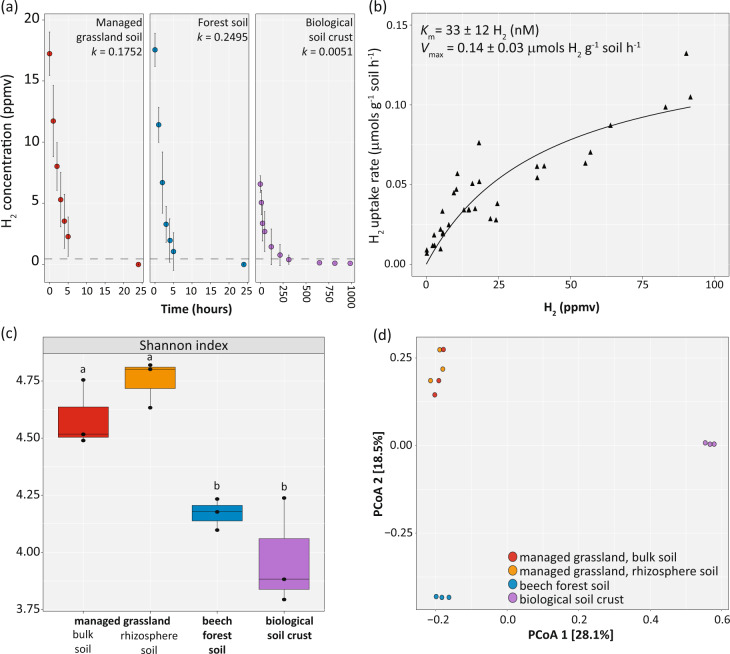
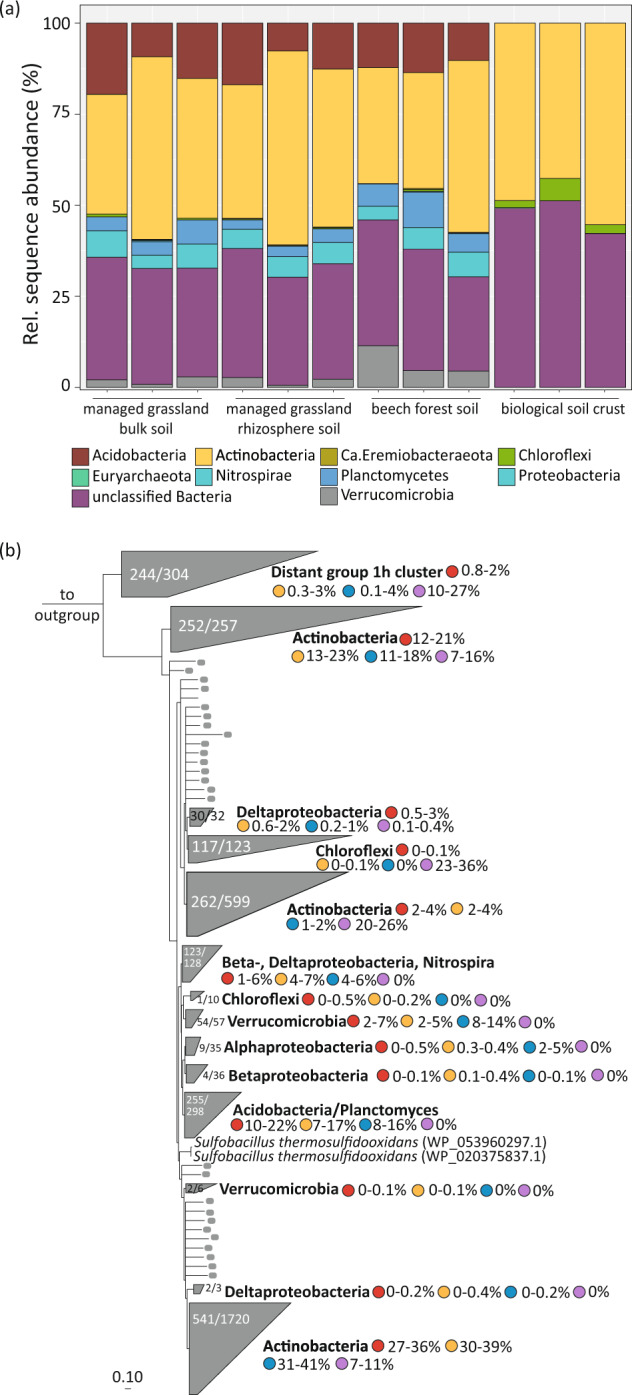
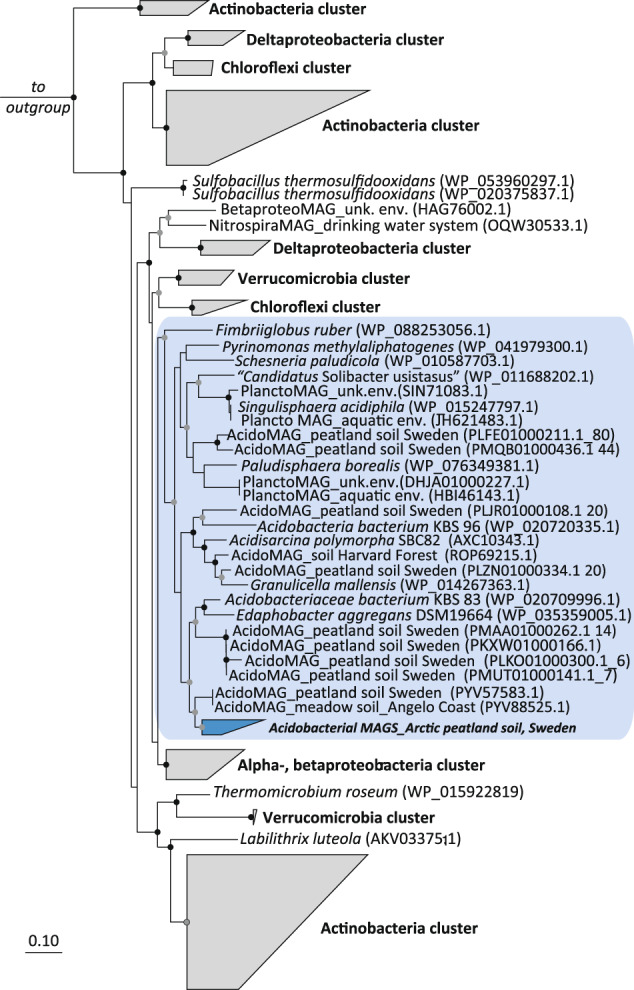
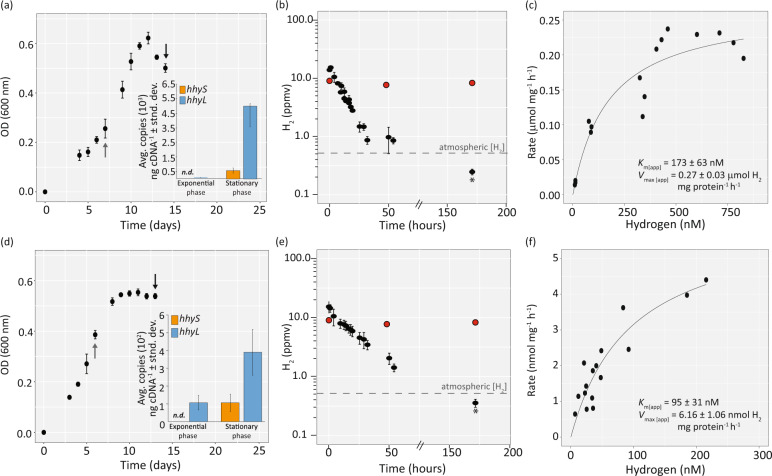
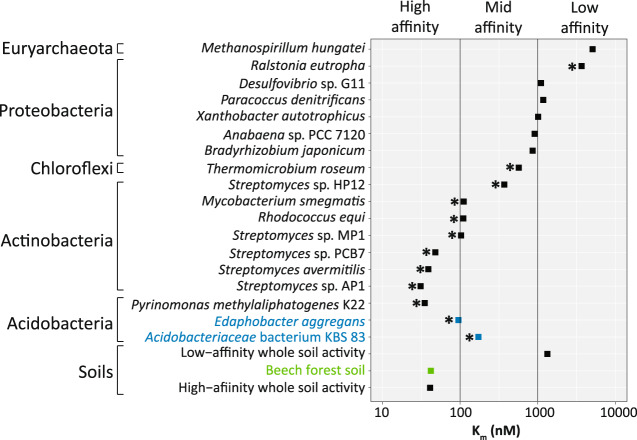
Significant rates of atmospheric dihydrogen (H2) consumption have been observed in temperate soils due to the activity of high-affinity enzymes, such as the group 1h [NiFe]-hydrogenase. We designed broadly inclusive primers targeting the large subunit gene (hhyL) of group 1h [NiFe]-hydrogenases for long-read sequencing to explore its taxonomic distribution across soils. This approach revealed a diverse collection of microorganisms harboring hhyL, including previously unknown groups and taxonomically not assignable sequences. Acidobacterial group 1h [NiFe]-hydrogenase genes were abundant and expressed in temperate soils. To support the participation of acidobacteria in H2 consumption, we studied two representative mesophilic soil acidobacteria, which expressed group 1h [NiFe]-hydrogenases and consumed atmospheric H2 during carbon starvation. This is the first time mesophilic acidobacteria, which are abundant in ubiquitous temperate soils, have been shown to oxidize H2 down to below atmospheric concentrations. As this physiology allows bacteria to survive periods of carbon starvation, it could explain the success of soil acidobacteria. With our long-read sequencing approach of group 1h [NiFe]-hydrogenase genes, we show that the ability to oxidize atmospheric levels of H2 is more widely distributed among soil bacteria than previously recognized and could represent a common mechanism enabling bacteria to persist during periods of carbon deprivation.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Rhee TS, Brenninkmeijer CAM, Röckmann T. The overwhelming role of soils in the global atmospheric hydrogen cycle. Atmos Chem Phys. 2006;6:1611–25.
-
- Downey NVS, Randerson JT, Eiler JM. Molecular hydrogen uptake by soils in forest, desert, and marsh ecosystems in California. J Geophys Res. 2008;113:G03037.
-
- Schmitt S, Hanselmann A, Wollschläger U, Hammer S, Levin I. Investigation of parameters controlling the soil sink of atmospheric molecular hydrogen. Tellus B Chem Phys Meter. 2009;61:416–23.
-
- Novelli PC, Lang PM, Masarie KA, Hurst DF, Myers R, Elkins JW. Molecular hydrogen in the troposphere: Global distribution and budget. J Geophys Res. 1999;104:30427–44.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
